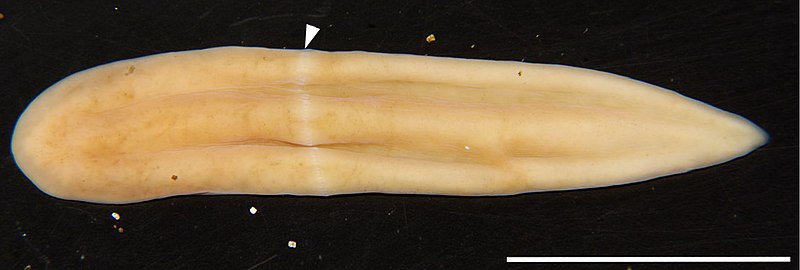
Quatro hepáticas já foram apresentadas aqui anteriormente, mas está mais do que na hora de falar sobre a hepática, a espécie que fez esse grupo de plantas receber esse nome comum. Marchantia polymorpha é seu nome científico e é geralmente chamada de hepática comum.
De ampla distribuição na região holártica (isto é, América do Norte, Europa e Norte da Ásia), a hepática-comum tem, como outras hepáticas, sua fase gametófito como a fase dominante. O gametófito é um talo achatado de até 10 cm de comprimento e 2 cm de largura, frequentemente verde, mas às vezes se tornando amarronzado ou roxo. A forma geral do talo lembra um fígado, de onde o nome hepática.
Hepática-comum crescendo entre alguns musgos. As pequenas estruturas circulares nos talos são conceptáculos. Foto de Krzysztof Ziarnek.*
O gametófito da hepática-comum pode ser macho ou fêmea. Ambas as plantas produzem estruturas em forma de guarda-chuva, os gametóforos, nos quais os gametas são produzidos. O gametóforo feminino tem uma estrutura em forma de estrela no topo, enquanto o gametóforo masculino tem um disco achatado, às vezes com margens lobadas. Quando os gametas masculinos estão maduros, são carregados com a água da chuva até os gametóforos femininos, onde fertilizam os gametas femininos. O zigoto se desenvolverá num esporófito, que cresce no lado inferior do gametóforo feminino, fazendo-o ficar “felpudo”. O esporófito, por sua vez, produzirá esporos que são liberados no ambiente e germinarão para originar novos gametófitos.
Gametóforos masculinos com forma de disco lobado.
Os gametófitos também podem se reproduzir assexuadamente produzindo gemas dentro de conceptáculos. As gemas são pequenas plantas em forma de lentilha que são liberadas quando gotas de água caem dentro dos conceptáculos.
Os gametóforos femininos com sua forma de estrela.
A hepática-comum é uma planta pioneira e coloniza solos expostos muito rapidamente. Após grandes incêndios florestais, ela pode rapidamente cobrir o solo da região afetada, assim prevenindo a erosão do solo. Por outro lado, sua dispersão rápida a faz uma erva daninha comum em jardins e estufas. Ela também é muito resistente a altas concentrações de chumbo no solo, de forma que regiões em que ela é muito abundante mas em que de outra forma poucas plantas crescem podem indicar um solo contaminado.
Um gametóforo feminino com os esporófitos felpudos crescendo dele.
Devido à sua aparência de fígado, ela foi usada historicamente para tratar afecções do fígado através da doutrina das assinaturas, que dizia que uma planta que se assemelha à forma de um órgão humano poderia ser usada para tratar doenças daquele órgão.
Close de uma gema, uma pequena estrutura para reprodução assexuada. Créditos ao usuário Des_Callaghan do Wikimedia Commons.*
Em décadas recentes, o interesse na hepática-comum como organismo modelo começou a crescer. Ela tem um ciclo de vida rápido, é facilmente cultivada e possui um genoma relativamente pequeno, o que a torna interessante para estudar vários aspectos biológicos, especialmente a evolução das plantas. No entanto seu genoma foi sequenciado apenas recentemente, em 2017, mas já mostrou que pode nos ajudar a entender a evolução das plantas para conquistarem a terra.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Bowman, J. L., Kohchi, T., Yamato, K. T., Jenkins, J., Shu, S., Ishizaki, K., … & Schmutz, J. (2017). Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell, 171(2), 287-304. https://doi.org/10.1016/j.cell.2017.09.030
Shimamura, M. (2016). Marchantia polymorpha: taxonomy, phylogeny and morphology of a model system. Plant and Cell Physiology, 57(2), 230-256. https://doi.org/10.1093/pcp/pcv192
De forma a tentar evitar predadores, muitas espécies desenvolvem poderosas toxinas que prejudicariam qualquer um tentando comê-las, às vezes até matando. No entanto os predadores podem contra-atacar desenvolvendo uma forte resistência às defesas da presa, às vezes a níveis espetaculares.
Milípedes, ou piolhos-de-cobra, são conhecidos como presas de sabor desagradável que evoluíram uma variedade de toxinas para deter predadores. Não obstante, algumas espécies encontraram maneiras de lidar com as defesas dos milípedes, tornando as pobres criaturas desesperadas pelo desenvolvimento de novas estratégias de sobrevivência.
Na América do Norte, besouros carabídeos do gênero Promecognathus são predadores especialistas em milípedes. A espécie Xystocheir dissecta, uma de suas principais presas, produz cianeto como defesa química. Cianeto é um composto muito tóxico para a maioria das formas de vida.
O milípede produtor de cianeto Xystocheir dissecta. Foto do usuário mhertel do iNaturalist.*
Num estudo recente, 18 espécies diferentes de besouros carabídeos foram expostas a cianeto de sódio (NaCN) em laboratório para terem sua resistência verificada. Enquanto a maioria das espécies sucumbiu em menos de 10 minutos quanto exposta a 15 mg de NaCN ou menos, três espécies pouco se lixaram mesmo para quantidades tão altas quanto 100 mg. Estas três espécies são Promecognathus crassus, P. laevissimus e Metrius contractus. Enquanto as duas espécies de Promecognathus se alimentam de Xystocheir dissecta, Metrius contractus não o faz.
Promecognathus laevissimus, o besouro-“eu comeria cianeto no café”. Foto de Eddie Dunbar.*
Em outro experimento, as espécies foram expostas de 100 mg de cianeto de potássio (KCN) por até duas horas. Enquanto M. contractus permaneceu ativo durante a primeira hora, todos os espécimes sucumbiram em menos de duas horas, mas, após 120 min, alguns espécimes de Promecognathus laevissius ainda estavam andando por aí como se nada estivesse acontecendo.
Metrius contractus, resistindo ao cianeto só de zoa. Foto do usuário tparkeressig do iNaturalist.*
Este estudo é a primeira evidência de predadores resistentes a cianeto. Enquanto este superpoder em P. laevissimus é facilmente explicado pelo seu comportamento predatório, a alta resistência de M. contractus ainda é um mistério, já que esta espécime não é especializada em milípedes, apesar de ser possível que os coma como comida alternativa, especialmente espécimes doentes ou machucados. Ambas as espécies, no entanto, são resistentes a quantidades de cianeto bem acima do que eles encontrariam em qualquer milípede. É realmente um superpoder.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referência:
Weary BP, Will KW (2020) The Millipede-Predation Behavior of Promecognathus and Exceptional Cyanide Tolerance in Promecognathus and Metrius (Coleoptera: Carabidae). Annals of the Entomological Society of America. https://doi.org/10.1093/aesa/saaa023
Bactérias são encontradas em quase todos os lugares do nosso planeta e são essenciais para a sobrevivência de todas as outras formas de vida, incluindo as fascinantes, e para alguns nojentas, baratas. Um gênero especial amigo das bactérias possui o apropriado nome Blattabacterium, cuja espécie mais bem conhecida é Blattabacterium cuenoti, a qual decidi chamar de “bactéria-de-barata-comum”.
Esta interessante espécie, como todas as outras espécies de Blattabacterium, é um endossimbionte obrigatório de baratas, o que significa que só pode existir dentro de células de baratas. Mais especificamente, a bactéria-de-barata-comum vive dentro das células dos corpos de gordura das baratas, isto é, seu tecido adiposo. Ela foi encontrada vivendo dentro de todas as espécies de baratas examinadas até hoje com exceção do gênero Nocticola. Ela também é encontrada dentro do cupim Mastotermes darwinensis porque, caso você ainda não saiba, cupins não são nada mais que baratas altamente especializadas. Assim, acredita-se que esta bactéria “infectou” pela primeira vez o ancestral de todas as baratas modernas cerca de 140 milhões de anos atrás e foi perdida em apenas duas linhagens, a de Nocticola e a dos cupins.
Células de Blattabacterium cuenoti mostradas em vermelho (acima) e cinza (abaixo). As áreas em ciano da imagem de baixo representam o núcleo das células da barata. Extraído de Sabree et al. (2009).
Apesar de muitas baratas serem generalistas, sendo capazes de comer quase tudo, a dieta principal de todas as espécies consiste em matéria vegetal em decomposição e este é um alimento relativamente pobre em nitrogênio. Então, de modo a aumentar o consumo de nitrogênio, as baratas armazenam ácido úrico, um produto comum do metabolismo de proteínas. A maioria dos animais, incluindo humanos, excreta ácido úrico em sua urina, mas baratas o armazenam em seu tecido adiposo. Assim, pensava-se originalmente que as bactérias-de-barata, por viverem perto das reservas de ácido úrico no tecido adiposo, pudessem usar ácido úrico diretamente como fonte de alimento, mas estudos descobriram que este não é o caso.
Quando necessário, as baratas liberam este ácido úrico e ele é quebrado em ureia ou amônia por bactérias vivendo no intestino. Depois disso, as bactérias-de-barata podem usar estes compostos e sintetizar glutamato, aminoácidos essenciais e vitaminas para a barata.
Como não podem usar ácido úrico diretamente, é um mistério por que as bactérias-de-barata vivem tão perto do lugar onde esta substância é armazenada. Uma sugestão é de que elas eram originalmente capazes de usar ácido úrico, mas perderam esta habilidade por redução do genoma.
As categorias de genes funcionais de Blattobacterium são muito similares àquelas de Blochmannia, uma bactéria endossimbionte de formigas-carpinteiras, as quais também se alimentam de matéria vegetal. Contudo Blochmannia é muito distantemente relacionada a Blattobacterium, sugerindo que seus genomas similares sejam o resultado de evolução convergente causada pelo estilo de vida similar.
Quando algo funciona, a natureza inventa mais de uma vez.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
López-Sanchez MJ, Neef A, Peretó J, Patiño-Navarrete R, Pignatelli M, Latorre A, Moya A (2009) Evolutionary Convergence and Nitrogen Metabolism in Blattabacterium strain Bge, Primary Endosymbiont of the Cockroach Blattella germanica. PLoS Genetics 5(11): e1000721. 10.1371/journal.pgen.1000721
Patiño-Navarrete R, Moya A, Latorre A, Peretó J (2013) Comparative Genomics of Blattabacterium cuenoti: The Frozen Legacy of an Ancient Endosymbiont Genome. Genome Biology and Evolution 5(2): 351–361. https://doi.org/10.1093/gbe/evt011
Sabree ZL, Kambhapati S, Moran NA (2009) Nitrogen recycling and nutritional provisioning by Blattabacterium, the cockroach endosymbiont. PNAS 106(46): 19521–19256. https://doi.org/10.1073/pnas.0907504106
Sete anos atrás eu discuti a posição filogenética de Acoelomorpha e seus parentes próximos, Xenoturbella, que juntos formam o clado Xenacoelomorpha. Sendo animais bilaterais muito simples que não possuem quase nenhuma das principais estruturas comuns à maioria dos outros bilaterais, sua posição filogenética exata é geralmente considerada basal dentro de Bilateria, mas a ideia de que são deuterostômios foi levantada depois que alguns estudos moleculares os agruparam dentro do clado Ambulacraria, que inclui equinodermos e hemicordados.
Pertencer aos deuterostômios significaria que Xenacoelomorpha sofreu uma gigantesca simplificação de sua anatomia. Lá em 2013, quando escrevi o outro artigo, isso estava causando bastante controvérsia, mas, pouco depois, novos estudos moleculares confirmaram a posição basal de Xenacoelomorpha e ficou meio que bem aceito que eles eram, de fato, o clado mais basal de Bilateria.
Uma versão simplificada da árvore da vida animal mostrando a posição incerta de Xenacoelomorpha. A posição de Placozoa e Ctenophora também não é muito clara.*
Mas uma vez problemático, sempre problemático.
Em 2019, um novo estudo que tentou antecipar os erros sistemáticos durante estudos de filogenia molecular, tal como atração de ramos longos, concluiu que a posição basal de Xenacoelomorpha é um artefato e que, quando se tenta minimizar os erros, sua posição como grupo irmão de Ambulacraria se torna clara. Contudo a árvore deles também sugere que Deuterostomia não seja monofilético, já que os cordados aparecem como grupo irmão de Protostomia e Xenacoelomorpha+Ambulacraria é o grupo mais basal, isto é, o grupo irmão do resto dos Bilateria. Contudo a ideia de Deuterostomia não ser monofilético é bem inesperada.
Como mencionei no meu último post, o principal problema em Xenacoelomorpha aparecer dentro de Deuterostomia tem a ver com sua supersimplificação. Eles não têm quase nada que um bilateral típico tem. O que os teria forçado a se tornarem tão simples?
Outro estudo recente sugere que, no caso de Xenoturbella ao menos, isso pode ser o resultado de seus hábitos de se enterrar num substrato macio. Eles comparam Xenoturbella a nudibrânquios, entre os quais algumas espécies possuem estilos de vida similares. Um desses nudibrânquios, Xenocratena, foi na verdade descoberto mais ou menos na mesma época que Xenoturbella vivendo no mesmo ambiente que ela. Eles possuem uma anatomia pedomórfica (mais simplificadas, “de bebê”) em comparação a outros nudibrânquios. Contudo ela não é de forma alguma tão simples quanto a de Xenoturbella.
Por outro lado, há outro gênero de nudibrânquios que é de fato supersimplificado, Pseudovermis, e ele vive enterrado em substrato macio também. Análises moleculares revelaram que Pseudovermis não é proximamente relacionado a Xenocratena, mas a Cumanotus, outro nudibrânquio que se enterra, o que sugere que esta simplificação ocorreu duas vezes entre os nudibrânquios.
Relações filogenéticas entre nudibrânqios. Veja Pseudovermis and Cumanotus a 2 horas e Xenocratena a 7 horas. Créditos a Martynov et al. (2020).*
Isso não é evidência de que Xenoturbella é um deuterostômio simplificado, mas e um bom argumento. Mas e quanto às simplificações em Acoelomorpha? Penso que se Xenoturbella não fosse proximamente relacionada a Acoelomorpha, eu estaria mais disposto a aceitar essa hipótese. Meu coração tende para a hipótese de Xenacoelomorpha basal, no entanto. Contudo, como qualquer cientista, aceitarei Xenacoelomorpha como deuterostômios se forem apresentadas evidências suficientes.
Xenoturbella é sempre o problema principal nesta equação. O sistema nervoso de Acoelomorpha, por exemplo, apesar de simplificado, tem meio que o padrão básico encontrado em todos os bilaterais e poderia ter evoluído do anel oral de um ancestral similar a um cnidário de acordo com algumas hipóteses. En Xenoturbella, no entanto, o sistema nervoso é muito mais estranho, sendo formando por uma rede difusa simples de neurônios abaixo da pele. Acho que abordar a organização do sistema nervoso em todos esses grupos é um bom tópico para outra postagem.
Se há uma coisa, na minha opinião, que faz a posição de Xenacoelomorpha dentro Deuterostomia ser algo convincente é o fato de muitas características de Deuterostomia parecerem ser mais primitivas dentro de Bilateria quando comparadas àquelas de Protostomia, de forma que a posição de Xenacoelomorpha entre os Deuterostomia é mais plausível que sua posição dentro de Protostomia (apesar de isso nem mesmo ser mais considerado uma possibilidade). Tendemos a pensar que deuterostômios são mais “derivados” simplesmente porque humanos são deuterostômios. Mas esta discussão também e assunto para outra postagem.
O que você acha? Você é time basal ou time deuterostômio?
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Cannon JT, Vellutini BC, Smith J, Ronquist F, Jonfelius U, Hejnol A (2016) Xenacoelomorpha is the sister group to Nephrozoa. Nature 530: 89–93. doi: 10.1038/nature16520
Jondelius U, Raikova OI, Martinez P (2019) Xenacoelomorpha, a Key Group to Understand Bilaterian Evolution: Morphological and Molecular Perspectives. In: Pontarotti P (ed) Evolution, Origin of Life, Concepts and Methods. Cham: Springer International Publishing, . pp. 287–315. doi: 10.1007/978-3-030-30363-1_14
Martynov A, Lundin K, Picton B, Fletcher K, Malmberg K, Korshunova T (2020) Multiple paedomorphic lineages of soft-substrate burrowing invertebrates: parallels in the origin of Xenocratena and Xenoturbella. PLoS ONE 15(1): e0227173. doi: 10.1371/journal.pone.0227173
Philippe H, Poustka AJ, Chiodin M, et al. (2019) Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria. Current Biology 29(11):1818–1826. doi: 10.1016/j.cub.2019.04.009
E com “todos nós” eu quero dizer nós, os eucariontes, os organismos com células complexas com núcleo, mitocôndrias e tal.
A forma como os organismos são classificados mudou enormemente ao longo dos dois últimos séculos, mas, durante as últimas décadas, ficou claro que temos três domínios da vida, os chamados Bacteria, Archaea e Eukarya. Apesar de a relação entre estes três domínios ter sido problemática no início, logo ficou claro que Eukarya e Archaea são mais proximamente relacionados entre si do que são com Bacteria.
Tanto Bacteria quanto Archaea são caracterizados pelas chamadas células procarióticas, nas quais não há núcleo delimitado e há somente um cromossomo circular (além de uma porção de anéis de genes menores chamados de plasmídeos). Eukarya, por outro lado, possui um núcleo cercado de uma membrana que inclui muitos cromossomos lineares. Tanto a estrutura da membrana celular quanto vários genes indicam que Archaea e Eukarya são proximamente relacionados, mas ainda era um mistério se ambos os grupos evoluíram de um ancestral comum e seriam, portanto, grupos irmãos, ou se eucariontes evoluíram diretamente de arqueias e seriam, portanto, arqueias altamente complexas.
As coisas passaram a apontar para a segunda hipótese após várias proteínas originalmente consideradas exclusivas de eucariontes (as chamadas Eukaryotic Signature Proteins, ESPs) foram encontradas em representantes do clado TACK de Archaea. Contudo clados diferentes dentro de TACK possuíam ESPs diferentes, então as coisas continuaram incertas.
Então em 2015 um novo grupo de arqueias foi descoberto no oceano Ártico entre a Noruega e a Groenlândia perto de um campo de fontes hidrotermais ativas chamadas Castelo de Loki (Spang et al. 2015). Chamado de Lokiarchaeota, este novo grupo de arqueias continha um grande número de ESPs, incluindo muitas das encontradas em diferentes linhagens TACK. Lokiarchaeota apareceu como grupo irmão de eucariontes em reconstruções filogenéticas e indicou que os eucariontes evoluíram, de fato, de arqueias, e aparentemente de arqueias mais complexas que as conhecidas até então. Este grupo foi baseado unicamente em um genoma incompleto encontrado nos sedimentos, já que o organismo em si não foi encontrado e não pôde ser cultivado para confirmar a estrutura de suas células.
Em 2016, outra linhagem de arqueias foi descoberta através de um genoma encontrado no estuário o rio White Oak na costa atlântica dos EUA (Seitz et al., 2016). Chamado Thorarchaeota, este clado revelou ser proximamente relacionado a Lokiarchaeota e, portanto, aos eucariontes.
Reconstrução das possíveis rotas metabólicas encontradas em Thorarchaeota com base no genes (quadrados brancos) encontrados no genoma torarqueótico. Créditos a Seitz et al. (2016).
Então em 2017 muitos novos genomas foram encontrados nos mesmos ambientes nos quais Lokiarchaeaota e Thorarchaeota haviam sido encontrados e em muitos outros (Zaremba-Niedzwiedzka et al., 2017). Eles incluíam dois novos grupos proximamente relacionados a esses dois, os quais foram chamados de Odinarchaeota e Heimdallarchaeota. O grupo inteiro recebeu o nome de “arqueias Asgard” e reconstruções filogenéticas puseram Eukarya dentro dele, com Heimdallarchaeota sendo o grupo irmão de Eukarya.
Mas perguntas e dúvidas logo surgiram. Ainda em 2017, um novo artigo (Da Cunha et al., 2017) questionou estes achados e levantou a hipótese de que as reconstruções filogenéticas que punham Asgard e Eukarya juntos eram um artefato causado por atração de ramos longos, um efeito colateral de reconstruções filogenéticas no qual espécies de evolução rápida forçam clados distantemente relacionados a colapsarem num único clado. A remoção de algumas arqueias de evolução rápida da análise foi suficiente para quebrar a relação Asgard-Eukarya. Visto que os genomas de Lokiarchaeota e outros Asgards foram reconstruídos a partir de DNA ambiental e não de células únicas, havia a possibilidade de as amostras estarem contaminadas com material de outros organismos. Os genes de proteínas usados nas análises também pareciam ter origens divergentes e podem ter sido adquiridos por transferência horizontal de genes, quando um gene é transferido de um organismo para outro por meios que não a reprodução, geralmente através de vírus.
Os autores originais do clado Asgard, que propuseram sua proximidade com Eukarya, rejeitaram o criticismo de Da Cunha et al. (2017) e afirmaram que estes usaram metodologia inadequada e que não havia evidência de contaminação em suas amostras (Spang et al., 2018).
(MDS, isso tá virando uma briga de verdade. Pega a pipoca!)
Da Cunha et al. (2018) responderam novamente mostrando mais evidência de contaminação e dizendo que Spang et al. deveriam mostrar evidência de metodologia inadequada se fosse o caso.
Estudos posteriores continuaram encontrando sequências de eucariontes em novas amostras de Asgard, o que reduziu a probabilidade de contaminação (Narrowe et al., 2018).
Fournier & Poole (2018) discutiram as implicações da proximidade de Eukarya com Asgard e propuseram uma classificação na qual Asgard não era mais membro de Archaea, mas formava um domínio novo, Eukaryomorpha, junto com Eukarya. Eles fizeram uma analogia com os mamíferos evoluindo de sinapsídeos e como sinapíseos costumavam ser vistos como répteis mesmo não estando dentro do clado Reptilia (Sauropsida). O mesmo seria o caso de Asgard. Apesar de terem “cara de Archaea”, eles não seriam arqueias de verdade.
Uma topologia hipotética de “arqueias verdadeiras”, Asgard e Eukarya de acordo com Fournier & Poole (2018).
Em um estudo publicado em dezembro passado, Williams et al. (2019) reanalisaram o caso usando mais dados e recuperaram novamente a proximidade de Asgard com Eukarya. Com este acúmulo de evidências, a hipótese de Eukarya surgindo de dentro de Archaea se tornou mais forte.
Aí agora, poucos dias atrás, finalmente conseguimos o que tanto queríamos. Um grupo de cientistas japoneses (Imachi et al., 2020) finalmente isolou um organismo Asgard e pôde fazer uma cultura em laboratório. Mas foi um trabalho difícil. A cultura cresceu muito devagar, com a fase lag (a fase em que as células se adaptam ao ambiente e crescem até dividirem) durando até 60 dias!
As criaturas estavam crescendo em uma cultura mista com uma bactéria do gênero Halodesulfovibrio e uma arqueia do gênero Methanogenium. As células Asgard foram chamadas de Candidatus Prometheoarcheum syntrophicum. Em taxonomia de procariontes, uma espécie nova recebe o status de Candidatus quando não é possível mantê-la numa cultura estável.
As células desta espécie de Asgard são cocoides, isto é, esféricas, e frequentemente apresentam vesículas na superfície ou longas protrusões na membrana que podem ou não se ramificarem. Estas protrusões não se conectam entre si nem a outras células, diferente de estruturas similares em outras arqueias. As células não parecem conter qualquer estrutura similar a organelas no interior, indo contra as expectativas. Asgard ainda não é a célula com cara de eucarionte que estávamos esperando!
Várias imagens de microscopia eletrônica de Candidatus Prometheoarchaeum syntrophicum. Vesículas mostradas em e, f e protrusões em g, h. Créditos a Imachi et al. (2020).
Graças à cultura desta espécie de Asgard, foi possível extrair seu genoma completo e confirmar o que era previamente conhecido de Asgard e baseado apenas em DNA ambiental. Isso confirmou a presença de 80 ESPs e, em uma análise filogenética, a espécie nova aparece como grupo irmão de Eukarya.
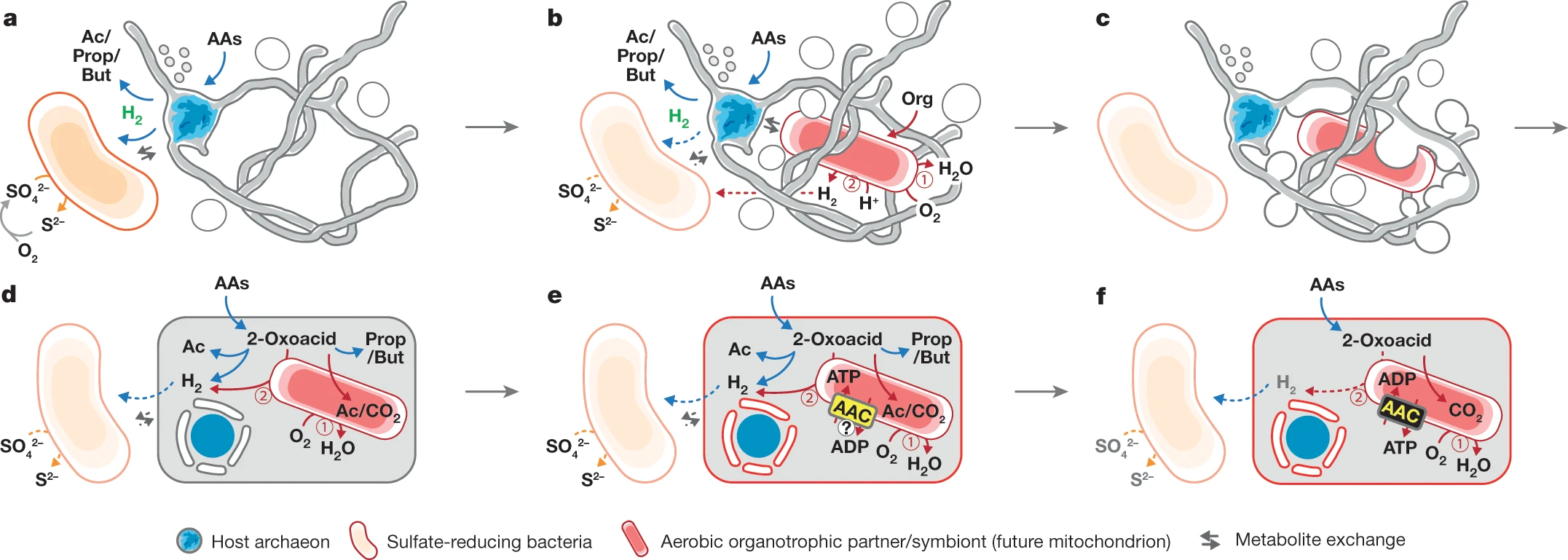
Candidatus Prometheoarcheum syntrophicum revelou ser anaeróbio e se alimentar de aminoácidos, quebrando-os em ácidos graxos e hidrogênio. Sua associação com os outros dois procariontes na cultura mista parece ser um tipo de mutualismo, com as três espécies se ajudando por transferência de hidrogênio de uma espécie para outra. Muitas perguntas sobre como um organismo assim se tornou uma célula eucarionte complexa ainda permanecem, mas ao menos temos mais algumas pistas sobre a aquisição da mitocôndria.
Hipótese da evolução da célula eucariótica baseada em uma relação mutualística entre uma arqueia do tipo Asgard e uma bactéria aeróbia. Créditos a Imachi et al. (2020).
A hipótese mais amplamente aceita era a de que uma célula eucarionte primitiva havia engolfado uma bactéria aeróbia através de fagocitose para comê-la, mas acabou retendo-a dentro de si. Contudo, vendo a cooperação de nossa arqueia Asgard com outros procariontes levanta a hipótese de que talvez o mutualismo entre a célula pro-eucarionte e as bactérias aeróbias começou quando ainda eram organismos separados.
Será que algum dia vamos achar de vez o “verdadeiro” proto-eucarionte? Vamos esperar pelos próximos episódios.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Da Cunha V, Gaia M, Gadelle D, Nasir A, Forterre P (2017) Lokiarchaea are close relatives of Euryarchaeota, not bridging the gap between prokaryotes and eukaryotes. PLOS Genetics 13(6): e1006810. doi: 10.1371/journal.pgen.1006810
Da Cunha V, Gaia M, Nasir A, Forterre P (2018) Asgard archaea do not close the debate about the universal tree of life topology. PLOS Genetics 14(3): e1007215. doi: 10.1371/journal.pgen.1007215
Imachi H, Nobu MK, Nakahara N et al. (2020) Isolation of an archaeon at the prokaryote–eukaryote interface. Nature. doi: 10.1038/s41586-019-1916-6
Narrowe AB, Spang A, Stairs CW, Caceres EF, Baker BJ, Miller SC, Ettema TJG (2018) Complex Evolutionary History of Translation Elongation Factor 2 and Diphthamide Biosynthesis in Archaea and Parabasalids. Genome Biology and Evolution 10: 2380–2393. doi: 10.1093/gbe/evy154
Seitz KW, Lazar CS, Hinrichs KU, Teske AP, Baker BJ (2016) Genomic reconstruction of a novel, deeply branched sediment archaeal plylum with pathways for acetogenesis and sulfur reduction. ISME Journal 10: 1696–1705. doi: 10.1038/ismej.2015.233
Spang A, Saw JH, Jørgensen SL, et al. (2015) Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521: 173–179. doi: 10.1038/nature14447
Spang A, Eme L, Saw JH, Caceres EF, Zaremba-Niedzwiedzka K, et al. (2018) Asgard archaea are the closest prokaryotic relatives of eukaryotes. PLOS Genetics 14(3): e1007080. doi: 10.1371/journal.pgen.1007080
Williams TA, Cox CJ, Foster PG, Szőllősi GJ, Embley TM (2019) Phylogenomics provides robust support for a two-domains tree of life. Nature Ecology & Evolution. doi: 10.1038/s41559-019-1040-x
Zaremba-Niedzwiedzka K, Caceres EF, Saw JH et al. (2017) Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541:353–358. doi: 10.1038/nature21031
Há uma coisa que eu deveria fazer mais frequentemente aqui, que é apresentar minha própria pesquisa para os leitores do blog, então hoje farei exatamente isso.
Como vocês devem saber, o grupo de organismos com que trabalho é a família Geoplanidae, comumente conhecidas como planárias terrestres. Aqui no Brasil, o gênero com mais espécies é Obama, do qual falei em posts anteriores. Este gênero se tornou consideravelmente famoso após uma de suas espécies, Obama nungara, se tornar invasora na Europa, o que chamou a atenção do público especialmente por causa do nome curioso do gênero, apesar de ele não ter nada a ver com o ex-presidente dos Estados Unidos.
Enfim, durante meu mestrado, ficou claro que espécies do gênero Obama se alimentam de invertebrados de corpo mole, especialmente lesmas e caracóis, apesar de algumas espécies também se alimentarem de minhocas ou mesmo outras planárias terrestres. Obama nungara, por exemplo, se alimenta dos três grupos, apesar de parecer preferir minhocas.
Um espécime de Obama anthropophila com suas sardas testiculares. Foto minha, Piter K. Boll.*
Um espécie de Obama comum em áreas urbanas do sul do Brasil é Obama anthropophila, cujo nome, significando “amante de humanos” é uma referência a este hábito precisamente. Esta espécie possui uma cor dorsal uniformemente marrom-escura, às vezes manchada pelos testículos maduros aparecendo como manchas escuras na primeira metade do corpo. A dieta desta espécie inclui caracóis, lesmas, nemertíneos e outras planárias terrestres, especialmente do gênero Luteostriata, e mais especialmente da espécie Luteostriata abundans, que ocorre muito comumente em áreas urbanas também.
Assista Obama anthropophila capturando diferentes presas.
Assim eu me perguntei… se O. anthropophila se alimenta de diferentes tipos de invertebrados, isso significa que cada tipo fornece nutrientes diferentes, de forma que uma dieta mista é necessária ou mais benéfica que uma composta de um só tipo de presa? Para averiguar isso, dividi espécimes adultos de O. anthropophila em três grupos, cada um recebendo uma dieta diferente:
Grupo Dela: alimentado apenas com a lesma Deroceras laeve Grupo Luab: alimentado apenas com a planária Luteostriata abundans Grupo Mixed: alimentado com ambas as presas de forma alternada
Os resultados não foram o que eu esperava. O grupo Mixed apresentou uma taxa de sobrevivência menor que os grupos com dieta de só uma presa. Outro aspecto interessante foi que o grupo Mixed apresentou uma tendência a passar o dia de receber uma lesma sem comer, comendo apenas as planárias após alguns dias recebendo as presas alternadamente.
Baseado na hipótese de que uma dieta mista é mais nutritiva, eu esperava que o grupo Mixed apresentasse um desempenho melhor, ou ao menos similar ao dos grupos de dieta única se não houvesse aumento no valor nutricional com uma presa adicional. Contudo os resultados indicam que uma dieta mista pode ser ruim para a planária, ao menos se o animal precisa comer algo diferente em cada refeição.
Não sabemos o que causa isso, mas minha ideia é de que talvez diferentes presas demandem diferentes processos metabólicos, como a produção de diferentes enzimas e tal, e ter que resetar seu metabolismo constantemente é muito custoso. Como resultado, o desempenho dos espécimes recebendo tal dieta diminuiu e os animais passam a evitar um dos tipos de alimento porque comer menos é menos perigoso que misturar comida.
Uma Obama anthropophila “grávida” prestes a pôr uma cápsula de ovos. Foto minha, Piter K. Boll.*
Outro aspecto interessante é que planárias recebendo uma dieta mista, mesmo morrendo mais cedo, punham cápsulas de ovos mais pesadas que os grupos de dieta única. Cápsulas de ovos mais pesadas geralmente significam mais embriões ou mais nutriente para os embriões, aumentando o sucesso reprodutivo. Mas como um animal morrendo pode ser melhor se reproduzindo que um animal saudável?
Bem, isso pode estar relacionado à hipótese do investimento terminal. Acredita-se, e é provado em alguns grupos, que um organismo pode aumentar seu investimento em reprodução quando eventos reprodutivos futuros não são esperados, isto é, quando um organismo “se dá conta” de que está prestes a morrer, ele põe todo seu esforço em se reproduzir para garantir que seus genes passem para gerações futuras.
Não podemos ter certeza de nada ainda. Mais estudos são necessários para entender melhor a relação de planárias terrestres com sua comida. O que podemos assegurar é que, assim como Obama nungara, Obama anthropophila pode acabar na Europa ou outro lugar logo porque sua dieta relativamente ampla e sua proximidade com humanos a fazem uma potencial nova espécie a ser acidentalmente espalhada pelo mundo.
Parasitas são um tipo especial de organismos que vivem sobre ou dentro de outras formas de vida, lentamente se alimentando delas, mas geralmente não as matando, apenas reduzindo sua condição física em algum grau. Essa é uma forma muito mais discreta de sobreviver do que matar ou arrancar pedaços inteiros com uma mordida, como predadores (tanto carnívoros quanto herbívoros) fazem. Todavia, diferente destas criaturas, parasitas costumam ser visto como desagradáveis e nojentos. Ainda assim, o parasitismo é a forma mais comum de conseguir alimento na natureza.
Quando apresentei o carrapato-de-rinoceronte numa Sexta Selvagem recente, mencionei o dilema causado por isso. Como o carrapato-de-rinoceronte é um parasita de rinocerontes, e rinocerontes estão ameaçados de extinção, uma prática comum para melhorar o sucesso reprodutivo destes mamíferos é remover seus carrapatos, mas isso pode acabar levando o carrapato-de-rinoceronte à extinção também.
Isso de fato já aconteceu com outros parasitas, como o piolho Coleocephalum californici, que era um parasita exclusivo do condor-da-Califórnia, Gymnogyps californianus. Para salvar o condor, uma prática comum entre veterinários trabalhando com os conservacionistas era tirar os piolhos das aves e, como resultado, este piolho está agora extinto. O prejuízo que o piolho causava ao condor era tão pouco, no entanto, que sua extinção não era nem um pouco necessária, sendo nada mais que um caso de negligência e falta de empatia com uma espécie pequena e não-carismática.
O piolho Rallicola (Aptericola) pilgrimi também desapareceu para sempre durante as campanhas de conservação para salvar seu hospedeiro, o kiwi-manchado-pequeno, Apteryx owenii, em outro trabalho falho.
Os esforços para salvar o kiwi-manchado-pequeno, Apteryxowenii, da extinção levou à extinção de seu piolho. Foto de Judi Lapsley Miller.*
A agora extinta Rallicola (Aptericola) pilgrimi. Créditos ao Museu da Nova Zelândia.**
Outro grupo de parasitas que está enfrentando a extinção são as pulgas. A espécie Xenopsylla nesiotes era endêmica da Ilha do Natal junto com seu hospedeiro, o rato-da-Ilha-do-Natal, Rattus macleari. A introdução do rato-preto, Rattus rattus, na ilha levou a um rápido declínio na população do rato-da-Ilha-do-Natal, que se tornou extinto no começo do século XX e, é claro, a pulga se extinguiu com ele. A pulga Acanthopsylla saphes provavelmente se tornou extinta também. Ela era parasita do quol-oriental, Dasyurus viverrinus, na Austrália continental. O quol-oriental hoje só é encontrado na Tasmânia, pois a população da Austrália continental se tornou extinta em meados do século XX. Contudo a pulga nunca foi encontrada em populações da Tasmânia, então provavelmente ela desapareceu na Austrália continental junto com a população local do hospedeiro.
A pulga-do-bobo-pequeno Ceratophyllus (Emmareus) fionnus. Foto de Olha Schedrina, Natural History Museum.*
Mas as coisas vêm mudando ultimamente e felizmente a visão sobre os parasitas está melhorando. Uma avaliação recente foi feita numa população de outra pulga, a pulga-do-bobo-pequeno, Ceratophyllus (Emmareus) fionnus. Esta pulga tem como hospedeiro específico o bobo-pequeno, Puffinus puffinus. Apesar de o bobo-pequeno estar longe de ser uma espécie ameaçada e ter muitas colônias ao longo da costa do Atlântico Norte, a pulga é endêmica da Ilha de Rùm, uma pequena ilha ao oeste da costa da Escócia. Devido à pequena população do hospedeiro nesta ilha, a pulga foi avaliada como vulnerável. Se a população do bobo-pequeno na ilha estivesse estável, as coisas estariam bem, mas, como você já deve imaginar, as coisas não estão bem. Assim como aconteceu na Ilha do Natal, o rato-preto também foi introduzido na Ilha de Rùm e se tornou um predador do bobo-pequeno, atacando seus ninhos.
O bobo-pequeno, Puffinus puffinus, é o único hospedeiro da pulga-do-bobo-pequeno. Foto de Martin Reith.**
Algumas ideias foram sugeridas para proteger a pulga da extinção. Uma delas é erradicar o rato-preto da ilha ou ao menos manejar sua população perto da colônia de bobos-pequenos. Outra proposta é translocar algumas pulgas para outra ilha para criar populações adicionais em outras colônias de bobos-pequenos.
Mas por que se dar ao trabalho de proteger parasitas? Bem, há razões o bastante. Primeiro, eles compreendem uma enorme parcela da biodiversidade do planeta e sua perda teria forte impacto sobre qualquer ecossistema. Segundo, eles são uma parte essencial da história evolutiva de seus hospedeiros e são, portanto, promotores de diversidade por seleção natural. Remover os parasitas de um hospedeiro eventualmente reduziria sua variabilidade genética e o deixaria mais vulnerável a novos parasitas. Devido à sua coevolução com o hospedeiro, parasitas também são uma fonte valiosa de informação sobre a ecologia e a história evolutiva do hospedeiro, ajudando-nos a conhecer a dinâmica de suas populações. Podemos até encontrar maneiras de lidar com nossos próprios parasitas ao estudar os parasitas de outras espécies, e parasitas são certamente algo que os humanos conseguiram coletar em grande número enquanto se espalhavam pelo globo.
Os parasitas podem ser incômodos, mas são necessários. Pode parecer que eles enfraquecem o hospedeiro a princípio, mas, a longo prazo, o que não te mata te fortalece.
Kwak ML (2018) Australia’s vanishing fleas (Insecta: Siphonaptera): a case study in methods for the assessment and conservation of threatened flea species. Journal of Insect Conservation 22(3–4): 545–550. doi: 10.1007/s10841-018-0083-7
Kwak ML, Heath ACG, Palma RL (2019) Saving the Manx Shearwater Flea Ceratophyllus (Emmareus) fionnus (Insecta: Siphonaptera): The Road to Developing a Recovery Plan for a Threatened Ectoparasite. Acta Parasitologica. doi: 10.2478/s11686-019-00119-8
Rózsa L, Vas Z (2015) Co-extinct and critically co-endangered species of parasitic lice, and conservation-induced extinction: should lice be reintroduced to their hosts? Oryx 49(1): 107–110. doi: 10.1017/S0030605313000628
Ao redor do mundo todo, muitas espécies animais se adaptaram a viver em ambientes de caverna, locais que são naturalmente desprovidos de luz, parcial ou inteiramente, e são, portanto, habitats pobres em nutrientes. A falta de luz faz com que seja impossível para plantas e outros organismos fotossintetizantes sobreviverem e, como resultado, há pouco alimento disponível para criaturas não fotossintetizantes. Estas criaturas dependem quase inteiramente de alimento que entra na caverna da superfície pela água ou por animais que se movem entre a superfície e as profundezas.
Devido à falta de luz nas cavernas, animais adaptados a este ambiente são geralmente desprovidos de olhos, pois ver não é possível de qualquer jeito, e brancos, porque não há necessidade de pigmentação na pele para se proteger da radiação ou para informar qualquer coisa visualmente. Por outro lado, sentidos químicos como olfato e paladar são frequentemente muito bem desenvolvidos.
Todas estas limitações tornam os ambientes de caverna relativamente pobres em espécies quando comparados a ambientes de superfície. Ou ao menos isso é o que parece à primeira vista. Há, é claro, muito menos espécies macroscópicas, como animais multicelulares, mas estes animais são eles mesmos um ambiente que pode abrigar uma vasta e desconhecida diversidade de microrganismos dentro deles.
Como vocês devem saber, a maioria dos, se não todos os, animais possui relações mutualísticas com microrganismos, especialmente bactérias, vivendo em seus intestinos. Esses microrganismos são essenciais para muitos processos digestivos, e muitos nutrientes que animais conseguem pela comida só podem ser obtidos com o auxílio desses amigos microscópicos. Os tipos de microrganismos no intestino de um animal estão diretamente relacionados com a dieta do animal. Por exemplo, herbívoros geralmente possuem uma alta diversidade de microrganismos que são capazes de degradar carboidratos, especialmente os complexos como a celulose.
Um estudo recente, conduzido na China com peixes do gênero Sinocyclocheilus, comparou a diversidade microbiana intestinal de diferentes espécies, incluindo algumas que vivem na superfície e algumas que são adaptadas a cavernas. Todas as espécie de Sinocyclocheilus parecem ser primariamente onívoras, mas espécies diferentes podem ter preferência por um tipo particular de comida, sendo mais carnívoras ou mais herbívoras.
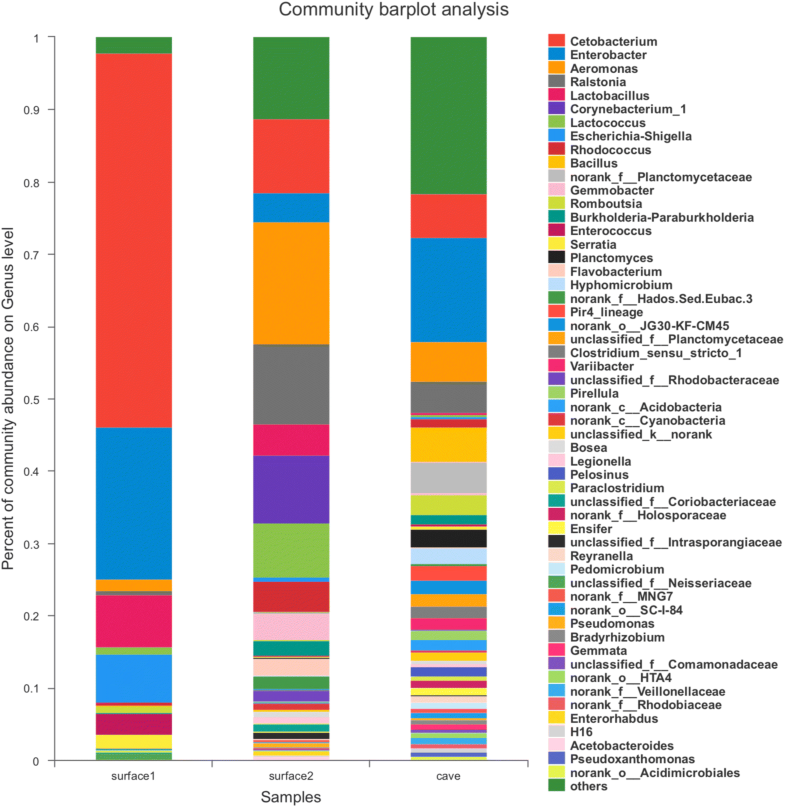
O estudo descobriu que espécies de Sinocyclocheilus de caverna possuem uma diversidade microbiana muito maior que espécies de superfície. Mas como isso pode ser possível se o número de recursos é limitado nas cavernas em comparação com a superfície? Bem, essa parece ser exatamente a razão.
Sinocyclocheilus microphthalmus, uma das espécies de caverna usada neste estudo. Foto extraída de Cool Goby Blog.
Como eu mencionei, espécie de Sinocyclocheilus são onívoras. Na superfície, elas possuem comida à vontade disponível e podem se dar ao luxo de escolher um tipo de comida preferido. Como resultado, sua microbiota intestinal é composta principalmente de espécies que auxiliam na digestão de um tipo específico de comida. Em cavernas, por outro lado, a comida é tão escassa que não se pode escolher e é preciso comer o que quer que esteja disponível. Isso inclui se alimentar de pequenas porções de diferentes tipos de alimento, incluindo outros animais que vivem na caverna e muitos tipos diferentes de resíduos animais e vegetais que chegam à caverna pela água. Assim, uma comunidade muito mais diversa de microrganismos intestinais é necessária para que a digestão seja eficiente.
Veja como o número de gêneros diferentes de bactérias é muito maior no grupo de caverna (direita) que em dois grupos da superfície (esquerda e centro). Imagem extraída de Chen et al. (2019)
Mais do que uma diversidade aumentada por si só, a comunidade de peixes de caverna também mostrou um grande número de bactérias que é capaz de neutralizar compostos tóxicos de vários tipos. A razão para isso ainda não é clara, mas há duas explicações possíveis que não são necessariamente mutualmente exclusivas. A primeira afirma que a água em cavernas é renovada numa taxa muito menor que águas superficiais, o que promove o acúmulo de todo tipo de substâncias, incluindo resíduos metabólicos das próprias espécies da caverna que podem ser tóxicos. A segunda hipótese é de maior preocupação e sugere que este aumento do número de bactérias capazes de degradar substâncias nocivas é um fenômeno recente causado pelo aumento de poluição da água por atividades humanas, o que está promovendo uma pressão seletiva em organismos de caverna.
A microbiota intestinal diversa de peixes de caverna é, portanto, uma estratégia desesperada mas inteligente de sobreviver num ambiente tão hostil. A natureza sempre encontra um meio.
Chen H, Li C, Liu T, Chen S, Xiao H (2019) A Metagenomic Study of Intestinal Microbial Diversity in Relation to Feeding Habits of Surface and Cave-Dwelling Sinocyclocheilus Species. Microbial Ecology. doi: 10.1007/s00248-019-01409-4
Todos estamos familiarizados com animais de muitas espécies que desenvolveram mecanismos interessantes para evitar serem comidos. Isso inclui, por exemplo, animais que se parecem com partes de plantas:
O famoso bicho-folha-gigante Phyllium giganteum. Foto de Bernard Dupont.**
e animais que se mesclam com o fundo:
Um chacal-da-África-Oriental, Canis mesomelas, na Savana. Consegue vê-lo? Foto de Nevit Dilmen.***
Também há animais que se parecem com outros animais não palatáveis ou perigosos para afastar os predadores:
A borboleta comestível vice-rei, Limenitis archippus (acima) imita a venenosa borboleta-monarca Danaus plexippus (abaixo). Créditos ao usuário do Wikimedia DRosenbach. Fotos de D. Gordon E. Robertson e Derek Ramsey.***
Mas nós raramente pensamos que as plantas também usam este tipo de mecanismo para evitar serem comidas. Há, no entanto, alguns casos registrados de comportamentos similares em plantas. Um caso é o da planta Corydalis benecincta, cujas folhas comumente possuem a cor amarronzada das rochas ao redor:
As folhas de Corydalis benecincta se parecem com as rochas encontradas em seu habitat natural. Foto de Yang Niu.
Recentemente, um estudo com plantas do gênero Amorphophallus encontrou outro caso interessante de imitação. Este gênero, que inclui o famoso jarro-titã, geralmente desenvolve uma única folha grande que em algumas espécies atinge o tamanho de uma pequena árvore ou de um arbusto. Uma folha gigante assim parece ser a refeição perfeita para um herbívoro, mas, para evitá-los, muitas espécies do gênero desenvolveram uma série de marcas ao longo do pecíolo da folha que se parecem com líquens ou cianobactérias.
Marcas semelhantes a cianobactérias no pecíolo de Amorphophallus gigas (A); Marcas semelhantes a cianobactérias + marcas semelhantes a líquens também em A. gigas (B); e marcas semelhantes a líquens em A. hewittii (C) e A. dactylifer (D). Extraído de Claudel et al. (2019).
Com esta imitação, os pecíolos, que são bem tenros, acabam se parecendo com um tronco duro e velho que não parece tão interessante para a maioria dos herbívoros. As marcas de líquens são tão bem representadas que podem até mesmo ser associadas com gêneros reais de líquens. Por exemplo, as marcas vistas nas figuras B e C acima se parecem com líquens do gênero Cryptothecia.
Líquens da espécie Cryptothecia striata, que parecem ser imitados pelas marcas em Amorphophallus gigas e A. hewittii. Foto de Jason Hollinger.*
Como e por que estas marcas evoluíram através das espécies de Amorphophallus ainda não é bem compreendido. Apesar da hipótese de que elas ajudam a planta a imitar um tronco de árvore, algumas espécies com folhas pequenas também possuem essas marcas, enquanto outras com folhas grandes não possuem qualquer marca ou as têm em padrões mais simples. O jarro-titã Amorphophallus titanum é um bom exemplo do último caso:
Amorphophallus titanum é a maior espécie de Amorphophallus, mas apresenta um padrão de líquen consideravelmente simples. Foto do usuário do flickr Bjorn S.**
Por muito tempo, as plantas foram consideradas organismos menos dinâmicos que os animais, mas em anos recentes nosso conhecimento sobre elas está aumentando e mostrando que na verdade elas são criaturas muito versáteis e desenvolveram estratégias igualmente criativas e complexas.
Claudel C, Lev-Yadun S, Hetterscheid W, & Schultz M 2019. Mimicry of lichens and cyanobacteria on tree-sized Amorphophallus petioles results in their masquerade as inedible tree trunks. Bot J Linn Soc 190: 192–214.
Canibalismo sexual é o ato de comer um parceiro sexual logo antes, durante ou logo depois da cópula. Apesar de ser um comportamento consideravelmente raro, sua ocorrência é muito popular entre o público em geral.
Quando canibalismo sexual ocorre, geralmente consiste na fêmea comendo o macho. Dois casos populares são os dos louva-a-deuses e das aranhas, especialmente a viúva-negra. Este fenômeno, ao menos entre viúvas-negras, é muito mais raro do que a maioria das pessoas pensa.
Louva-a-deus-fêmea comento um macho saboroso. Foto de Oliver Koemmerling.*
Apesar de às vezes o canibalismo sexual ocorrer porque um dos parceiros confunde o outro com comida, em muitas espécies é uma estratégia selecionada evolutivamente para assegurar que a fêmea coma o suficiente para a prole se desenvolver adequadamente. Pode parecer horrível do ponto de vista humano, especialmente se pensarmos da perspectiva do macho, mas temos que lembrar que passar os genes para a próxima geração é o principal propósito da maioria dos organismos e, se o macho é bem-sucedido em fertilizar os ovos da fêmea, sua vida serviu seu propósito e ele pode morrer feliz.
Canibalismo sexual é, claro, quase exclusivamente observado entre predadores, o que é um tanto óbvio. E, como eu disse acima, ele é comumente realizado pela fêmea. Um grupo que é famoso por suas espécies com fêmeas empoderadas é a ordem de insetos Hymenoptera, que inclui abelhas, formigas, vespas, moscas-serra, entre outros. Visto que muitos himenópteros apresentam algum grau de socialidade, na qual sociedades são compostas quase exclusivamente por fêmeas, e os machos são gerados apenas para a reprodução, é curioso que canibalismo sexual nunca tenha sido registrado neste grupo… até agora.
Um estudo recentemente publicado examinou o comportamento de acasalamento de uma pequena vespa parasitoide, Gonatopus chilensis. Esta espécie pertence à família Dryinidae, da qual todas as espécies põem os ovos em insetos da subordem Auchenorrhyncha, que inclui cigarras, cigarrinhas, jequitiranaboias, entre outros. As larvas, após eclodirem do ovo, se alimentam do hospedeiro. Fêmeas adultas de vespas driinídeas também são predadores vorazes e se alimentam das mesmas espécies das quais se alimentaram como larvas.
Macho de Gonatopus chilensis (esquerda) inseminando uma fêmea (a), e fêmea comendo um macho (b e c). Extraído de Vira & Espinosa (2019).*
Após a cópula, fêmeas de G. chilensis foram frequentemente observadas tentando capturar os machos da mesmo forma como capturam as presas. Contudo apenas em uma ocasião a fêmea foi bem-sucedida e capturar o macho e comeu seu gáster (a porção grande e arredondada que forma a maior parte do abdome em vespas). Como apenas um evento de canibalismo foi observado, ele pode ser um fenômeno raro nesta espécie, mas como várias tentativas de capturar o macho foram vistas, parece que comer o macho é uma ideia interessante para as fêmeas.
Este é o primeiro caso conhecido de canibalismo sexual em himenópteros e, portanto, um registro importante que aumentou o número de grupos nos quais se sabe que este comportamento ocorre.
Virla EG, Espinosa MS (2019) Observations on the mating behavior of a dryinid and first record of sexual cannibalism in the hymenoptera. Acta Ethologica. doi: 10.1007/s10211-019-00315-9






 Esta obra está licenciada sob uma Licença Creative Commons de Atribuição e Compartilhamento Igual 4.0 Internacional.
Esta obra está licenciada sob uma Licença Creative Commons de Atribuição e Compartilhamento Igual 4.0 Internacional.