Infinitas formas de beleza existem entre as pequenas criaturinhas que frequentemente nos cercam. Ácaros, que são tão onipresents, contêm várias belezas negligenciadas. Uma delas é o camaradinha de hoje, Eatoniana plumipes, que eu decidi batizar de ácaro-pé-de-pluma-do-Mediterrâneo.
Ácaro-pé-de-pluma-do-Mediterrâneo fotografado da Espanha por Simon Oliver*.
Adultos do ácaro-pé-de-pluma-do-Mediterrâneo são consideravelmente grandes para um ácaro, medindo uns poucos milímetros de comprimento, geralmente mais de 1 cm quando as pernas são consideradas. Eles são marrom-avermelhados, mais claros nas pernas e outros apêndices, e suas pernas traseiras são muito mais longas que as outras e possuem um tufo de pelos pretos longos que as faz parecer uma pluma, por isso o nome pé-de-pluma. Como sugerido pelo nome comum, a espécie é encontrada nos arredores do Mediterrâneo, incluindo o sul da Europa, norte da África, Turquia e o Oriente Médio.
Apesar de de ser um ácaro grande e consideravelmente bonito, muito pouco é conhecido sobre a história de vida do ácaro-pé-de-pluma-do-Mediterrâneo. Ele pertence a um grupo de ácaros que são predadores quando adultos, mas parasitas quando larvas. As larvas eclodem de ovos vermelhos postos pelas fêmeas no ambiente e são, é claro, muito menores que os adultos. Elas também têm só três pares de pernas, não quatro como os adultos, e não possuem as plumas características vistas nos adultos.
Alguns ovos do ácaro-pé-de-pluma-do-Mediterrâneo e uma larva recém-eclodida. Extraído de Mąkol & Sevsay (2015).
Quase nada é conhecido sobre os hábitos alimentares desta espécie. Gafanhotos estão entre os hospedeiros identificados das larvas, mas é provável que outros artrópodes sejam parasitados também. As larvas se prendem às pernas dos hospedeiros e se alimentam ali, sugando sua hemolinfa (o “sangue” dos artrópodes). Não consegui encontrar informação nenhuma sobre quais espécies servem de presa para os adultos.
Apesar de não sabermos quase nada sobre a ecologia do ácaro-pé-de-pluma-do-Mediterrâneo, ainda podemos apreciar sua beleza, e essa espécie certamente desempanha algum papel fundamental em seu ecossistema.
Se você vive ao redor do Mediterrâneo, já viu algum deles por aí? Comente aí!
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
Humanos podem ser infectados por um grande número de diferentes parasitas, mas nenhum parasita possui um impacto tão grande na nossa espécie como aqueles do gênero Plasmodium, que causam a doença conhecida como malária. Várias espécies diferentes de Plasmodium infectam humanos, mas hoje vamos falar da mais mortal delas, Plasmodium falciparum, que eu decidi chamar de plasmódio-mortal.
O gênero Plasmodium pertence ao filo Apicomplexa, um grupo de protistas exclusivamente parasitas que evoluíram de algas de vida livre. Além de Plasmodium, outro importante apicomplexo que infecta humanos é o toxo, que causa toxoplasmose e já apareceu como um camaradinha aqui alguns anos atrás.
O ciclo de vida do plasmódio-mortal é muito complexo e inclui dois hospedeiros, um humano e um mosquito do gênero Anopheles. Quando um mosquito infectado pica um humano, ele libera entre 20 e 200 esporozoítos, que são um dos estágios de vida do plasmódio-mortal. Os esporozoítos são células alongadas que deslizam pela corrente sanguínea do humano infectado até atingirem o fígado. Eles usam seu complexo apical, uma estrutura formada de várias glândulas e organelas, para penetrar as células do fígado. O complexo apical é perdido no processo.
Ciclo de vida do plasmódio-mortal. Créditos a La Roche Lab, UC Riverside.**
Dentro das células do fígado, o esporozoíto se transforma num trofozoíto, que vive dentro de um vacúolo e começa a crescer e sofrer mitose e meiose sem divisão celular até se tornar uma célula única com dezenas de milhares de núcleos chamada esquizonte. O esquizonte eventualmente arrebenta em dezenas de milhares de células haploides pequenas chamadas merozoítos, que entram novamente na corrente sanguínea. Dotados de novos complexos apicais, os merozoítos agora o usam para entrar em glóbulos vermelhos e se transformam novamente em trofozoítos, que agora são haploides. Esse trofozoíto haploide que infecta glóbulos vermelhos cresce de novo e passa por mitose para formar um novo grande esquizonte que eventualmente vai arrebentar de novo em novos merozoítos que entram na corrente sanguínea mais uma vez e infectam novos glóbulos vermelhos, recomeçando o ciclo. Dentro dos glóbulos vermelhos, os trofozoítos se alimentam de hemoglobina e convertem parte dela num pigmento granular insolúvel chamado hemozoína.
Um trofozoíto (esquerda) e um esquizonte (centro, abaixo) em glóbulos vermelhos.
Todas as células de plasmódio infectando glóbulos vermelhos possuem seu ciclo em sincronia, de forma que todos os merozoítos são liberados na corrente sanguínea e infectam novas células simultaneamente. O ciclo dentro dos glóbulos vermelhos dura cerca de 48 horas e está relacionado com a típica manifestação cíclica de febre e calafrios causada pela malária falciparum.
Eventualmente alguns merozoítos se transformam em formas sexuais, os gametócitos masculino e feminino, conhecidos como microgametócito e macrogametócito, respectivamente. Elas levam entre uma e duas semanas para atingir a maturidade. Após atingirem esse estágio, se o humano infectado é mordido por um mosquito, os gametócitos são ingeridos e, dentro do intestino do mosquito, passam por mudanças morfológicas. O macrogametócito cresce para uma forma maior e esférica chamada macrogameta. O microgametócito sofre divisão celular em 15 minutos e se dividi em oito microgametas flagelados. Um microgameta fertiliza um macrogameta, o que origina um zigoto. O zigoto se desenvolve em uma célula móvel chamada oocineto que penetra os tecidos do mosquito e se transforma num oocisto imóvel. O oocisto cresce e sofre divisão nuclear até se tornar uma grande célula multinucleada chamada esporoblasto. O esporoblasto arrebenta em milhares de esporozoítos, que migram até as glândulas salivares do mosquito e, quando o mosquito pica outro humano, são liberados na corrente sanguínea, recomeçando o ciclo.
Um macrogametócito (esquerda) e um microgametócito (direita) entre glóbulos vermelhos.
Os principais efeitos deletérios causados pela malária em humanos ocorrem devido à destruição massiva dos glóbulos vermelhos e a produção de hemozoína, que é tóxica aos tecidos e começa a se acumular em diversos órgãos, prejudicando sua função. Apesar de várias espécies de Plasmodium causarem malária em humanos, cerca de 50% de todas as mortes por malária são causadas apenas pelo plasmódio-mortal, fazendo dele o mais mortal de todos os parasitas em humanos.
O plasmódio-mortal aparentemente evoluiu cerca de 10 mil anos atrás na África a partir de uma espécie muito similar que infecta gorilas. Atualmente, apesar de a maioria das infecções e mortes ainda ocorrerem na África subsaariana, o plasmódio-mortal se espalhou ao redor do mundo e também é prevalente próximo ao equador na América do Sul e na Ásia.
Devido à alta letalidade da malária falciparum a humanos, a doença causa uma forte pressão seletiva em populações humanas em áreas onde o parasita é comum. Assim, apesar de alguns dizerem que a seleção natural não afeta mais humanos, o plasmódio-mortal está aí para refutar esse argumento.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Lambros, C., & Vanderberg, J. P. (1979). Synchronization of Plasmodium falciparum erythrocytic stages in culture. The Journal of parasitology, 418-420.
Maier, A. G., Matuschewski, K., Zhang, M., & Rug, M. (2019). Plasmodium falciparum. Trends in parasitology, 35(6), 481-482. https://doi.org/10.1016/j.pt.2018.11.010
Talman, A. M., Domarle, O., McKenzie, F. E., Ariey, F., & Robert, V. (2004). Gametocytogenesis: the puberty of Plasmodium falciparum. Malaria journal, 3(1), 1-14. https://doi.org/10.1186/1475-2875-3-24
Plantas são o organismo padrão quando pensamos em fotossíntese, mas várias espécies na verdade perderam a habilidade de sintetizar seu próprio alimento usando luz e se tornaram completamente heterótrofas. Como resultado, tais plantas sobrevivem parasitando outras plantas e se alimentando de sua seiva. Provavelmente a mais famosa espécie de planta heterótrofa é a Rafflesia arnoldii, a flor-cadáver, que possui as maiores flores de qualquer planta e foi uma das primeiras Sexta Selvagens quase 10 anos atrás. Mas hoje eu vou apresentar outra planta heterótrofa bem distantemente relacionada a essa, Balanophora fungosa, conhecida como a raiz-fungo.
Essa espécie ocorre no sudeste da Ásia e na Austrália, onde vive no solo e parasita as raízes de várias plantas diferentes. Como em outras plantas heterótrofas, a raiz-fungo passa a maior parte de sua vida no subsolo como nada além de um sistema de raízes e rizomas presos à planta hospedeira. Ela só é visível na superfície quando produz suas flores, as quais, como a flor gigante da flor-cadáver, são bem incomuns.
Um grupo de inflorescências saindo do solo na Nova Caledônia. Podemos ver as brácteas pálidas e a clave aveludada de flores femininas rodeada por flores masculinas maiores na base. Foto do usuário juju98 do iNaturalist.*
As flores ocorrem em inflorescências que na verdade são até fofas. A cor geral varia de creme pálido, quase branco, a rosa. A base da inflorescência possui várias brácteas (folhas modificadas associadas a flores) que possuem a mesma cor creme-pálida a rosa, sem qualquer sinal de verde. A parte superior possui uma estrutura em forma de clava de aspecto aveludado formada por centenas de minúsculas flores femininas. Circundando a base da clava há algumas flores masculinas, que são muito maiores que as femininas, mas ainda muito pequenas. A inflorescência como um todo se parece com alguns cogumelos como as bufas, sendo essa a provável razão do nome comum raiz-fungo.
Um close numa inflorescência na Austrália onde podemos ver as flores masculinas e femininas em mais detalhes. Foto de Aaron Bean.*
Não consegui encontrar muitos detalhes sobre o ciclo de vida desse parasita tão fofinho, mas a raiz-fungo parece ser polinizada por uma enorme quantidade de animais, incluindo vários tipos de insetos, aracnídeos e até pequenos vertebrados, os quais podem ser atraídos para se alimentar do pólen ou néctar ou talvez atraídos pelo incomum cheiro que as flores produzem. O cheiro é diferente da fragrância doce da maioria das flores, mas também não é um cheiro desagradável de carniça como o da flor-cadáver, do jarro-titã e de muitas outras plantas. Na verdade, é dito que as flores possuem cheiro de rato. Será que esse cheiro engana pequenos mamíferos que pensam que se trata de um membro em época reprodutiva da própria espécie, assim como algumas orquídeas fazem imitando a forma e o cheiro de abelhas fêmeas? Essa é uma possibilidade, mas, na verdade, a maioria dos mamíferos e aves que visitam as flores são nectarívoros e provavelmente estão apenas em busca do delicioso néctar.
De qualquer forma há muito que ainda não conhecemos dessa tão incomum e adorável planta com cara de fungo.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
Os nematódeos são frequentemente chamados de vermes cilíndricos por seu corpo em forma de tubo. Alguns, no entanto, deixaram de ter esse formato típico, ao menos em parte de sua vida. Entre estes se encontram os nematódeos dos cistos, que formam o gênero Heterodera, um dos quais é Heterodera glycines, conhecido como nematódeo-do-cisto-da-soja.
Como você deve ter adivinhado pelo nome da espécie incluir “soja”, esse é um parasita da soja e é, de fato, uma das pragas mais devastadoras a atacar plantações de soja. Esses vermes passam a maior parte da vida dentro das raízes da soja, alimentando-se de de seus tecidos.
O aspecto típico dos cistos nas raízes de uma planta infectada.
Os adulto geralmente medem menos de 1 mm de comprimento, mas os machos e as fêmeas são bem diferentes em forma e comportamento. Os machos são mais longos que as fêmeas, mas muito finos, tendo o formato típico de nematódeos. Uma vez atingindo a fase adulta, eles deixam as raízes da soja e vão à procura de fêmeas para acasalarem. As fêmeas, por outro lado, possuem um corpo em formato de limão, quase esférico. Elas nunca deixam as raízes da soja mas, à medida que se tornam sexualmente maduras, seus corpos incham de ovos e eventualmente sua extremidade posterior arrebenta para fora das raízes, aparecendo como pequenos cistos brancos visíveis a olho nu.
Uma fêmea cheia de ovos. Créditos a Agroscope FAL Reckenholz , Swiss Federal Research Station for Agroecology and Agriculture.*
Preenchida com 200 a 400 ovos, a fêmea morre e se transforma num cisto escuro. Os ovos permanecem dentro do corpo morto da fêmea até as condições ambientais estarem adequadas. Durante esse período, o embrião se desenvolve de um estágio Jovem 1 (J1) num Jovem 2 (J2). O J2 deixa o ovo e procura uma nova raiz de soja para penetrar, onde então continua a se desenvolver até atingir a fase adulta.
Uma foto de microscopia eletrônica de varredura de um ovo e um jovem do nematódeo-do-cisto-da-soja.
Os primeiros sintomas da infecção na planta são bem sutis, como um leve amarelamento das folhas, o que é frequentemente confundindo com carência de nutrientes. O problema é frequentemente percebido somente na colheita, quando ocorrem perdas de até 30% na produção.
Ainda não há formas eficazes de eliminar o nematódeo-do-cisto-da-soja de áreas infectadas. Sabe-se que alguns fungos parasitas infectam o nematódeo e controlam sua população, e rotação de culturas é outra estratégia para reduzir a carga de infecção, já que plantar uma planta diferente por dois anos consecutivos mostra uma redução significativa no número de ovos viáveis no solo.
Enquanto algumas variedades de soja são resistentes à infecção, há também diferentes raças do nematódeo-do-cisto-da-soja que apresentam diferentes graus de virulência, e novas raças, frequentemente mais virulentas, estão surgindo constantemente para conseguir vencer a resistência desenvolvida pelas plantas. É a evolução sendo incentivada bem nas nossas fazendas.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
Yan, G., & Baidoo, R. (2018). Current research status of Heterodera glycines resistance and its implication on soybean breeding. Engineering, 4(4), 534-541. https://doi.org/10.1016/j.eng.2018.07.009
É hora de apresentarmos outra bactéria e por que não uma que nos ama mesmo nós a odiando? Chlamydia trichomatis é a camarada de hoje, uma bactéria meio incômoda que infecta humanos e às vezes pode causar problemas sérios.
A clamídia-humana pertence ao filo de bactérias conhecido como Chlamydiae, que é simplesmente o plural do nome da bactéria. Todas as clamídias parecem ser endossimbiontes obrigatórios de células eucarióticas, seja como parasitas ou em uma relação mutualística. A clamídia-humana, claro, é do primeiro tipo. Esta espécie é um parasita exclusivo de humanos e aparentemente não consegue infectar células de nenhuma outra espécie.
O ciclo de vida da clamídia-humana é similar ao de outras espécies de clamídia. Ela tem duas formas distintas conhecidas como corpos elementares e corpos reticulados. Os corps elementares são uma forma semelhante a um esporo medindo de 200 a 400 nanômetros de diâmetro. Eles possuem uma parede celular bem rígida e são capazes de sobreviver fora da célula hospedeira. Quando um corpo elementar entra em contato com uma célula hospedeira humana, principalmente células de mucosas, ele faz a célula hospedeira criar um vacúolo dentro do qual permanece. Esse vacúolo é conhecido como inclusão.
Inclusões de clamídia (as grandes bolhas na célula central) vistas sob o microscópio.
Com a inclusão, o corpo elementar muda para um corpo reticular metabolicamente ativo, o qual mede entre 600 e 1500 nanômetros. O corpo reticulado é capaz de modificar a inclusão para um ambiente mais adequado e começa a se replicar muito rapidamente até encher a célula hospedeira com bactérias em até 72 horas. Nesse ponto, os corpos reticulados mudam de volta para corpos elementares e fazem a célula hospedeira estourar e liberá-los, de forma que eles podem se espalhar para outras células e infectá-las também.
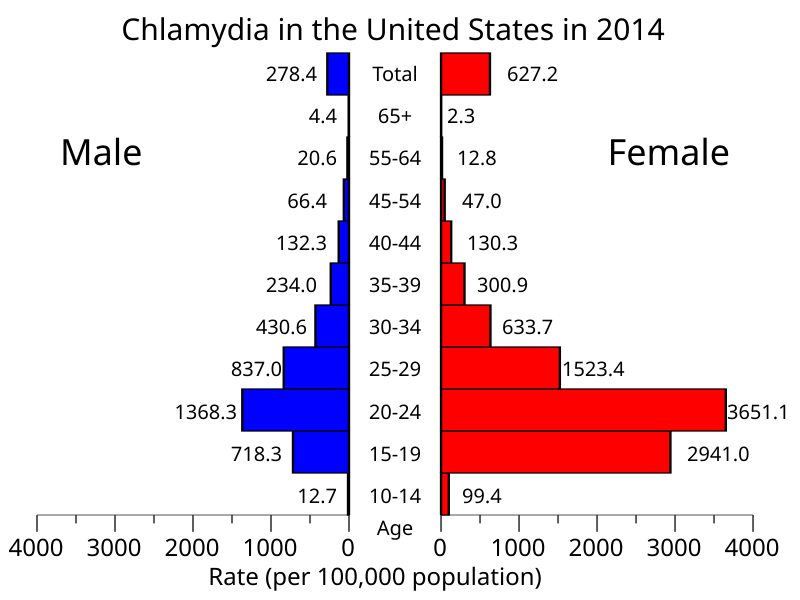
A clamídia-humana pode infectar muitas partes do corpo humano, mas as áreas mais comumente afetadas são a uretra e a vagina e sua transmissão entre humanos ocorre principalmente por relações sexuais, já que a pessoa infectada pode ter corpos elementares em seus fluidos, como esperma e fluido vaginal. De fato, clamídia é a infecção sexualmente transmissível mais comum no mundo, com cerca de 4.2% de todas as mulheres e 2.7% de todos os homens tendo a bactéria em seu corpo.
Muitos casos de infecção por clamídia passam sem ser diagnosticados porque às vezes a infecção não causa sintomas ou eles levam tempo demais para aparecer. Quando ela infecta a vagina e o colo do útero, os sintomas são raros no início, mas, à medida que a infecção se espalha, ela pode infectar o resto do sistema reprodutor e causar a chamada doença inflamatória pélvica, o que pode levar a esterilidade. Alguns dos raros sintomas de infecção vaiginal são dor durante a relação sexual e sangramento vaginal. Na uretra os sintomas são mais comuns e incluem dor ou uma sensação de queimação ao urinar e eventualmente um corrimento incomum. Os sintomas são bem similares àqueles de gonorreia.
Além do trato urogenital, a clamídia-humana pode infectar muitos outros locais, como o reto e a cavidade oral através de sexo anal e oral, respectivamente. Outra área comumente infectada são os olhos, com 19% de todos os casos de conjuntivite sendo causados pela clamídia-humana. Se não tratada adequadaemente, essa conjuntivite evolui para uma condição crônica conhecida como tracoma que frequentemente causa cegueira. Os olhos se tornam infectados por contato direto das mãos ou objetos (como toalhas) infectados com os olhos. A bactéria também pode ser transmitida por moscas enquanto elas andam por aí lambendo fluidos corporais humanos.
Infecção por clamídia de acordo com idade e sexo nos Estados Unidos.
Clamídia é frequentemente tratada com antibióticos como azitromicina. A prevenção inclui higiene adequada, práticas sexuais seguras e testagem regular em humanos sexualmente ativos, já que identificar a infecção cedo reduz seu dano e previne que se espalhe para outros. Na última década tem crescido o interesse em desenvolver uma vacina contra clamídia. Um problema é o fato de que a resposta imune contra essa bactéria parece ser muito complexa. Contudo testes preliminares com uma vacina candidata levou a resultados promissores, então há esperança!
Apesar de ser um pé no saco (ou, mais comumente, um ardor no saco) para humanos, a clamídia-humana é ao mesmo tempo um organismo fascinante como qualquer outra forma de vida da Terra (com exceção do Bostonauro e a escória que o elegeu). Ela possui um genoma consideravelmente pequeno, com somente cerca de 900 genes. Muitos genes metabólicos essenciais estão ausentes e acredita-se que a bactéria usa os do hospedeiro.
Um parasita específico de humanos, a clamídia-humana parece ter se tornado uma linhagem separada de outras espécies de clamídia cerca de 9 milhões de anos atrás. Isso significa que ela está conosco desde antes mesmo de sermos humanos.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Brunham, R. C., & Rey-Ladino, J. (2005). Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nature reviews immunology, 5(2), 149-161. https://doi.org/10.1038/nri1551
Manavi, K. (2006). A review on infection with Chlamydia trachomatis. Best Practice & Research Clinical Obstetrics & Gynaecology, 20(6), 941-951. https://doi.org/10.1016/j.bpobgyn.2006.06.003
Parasitas são quase tão velhos quanto a própria vida e podem ser um incômodo ameno a moderado ao hospedeiro na maior parte do tempo. Contudo ao domesticarmos muitas espécies, nós humanos ajudamos muitos parasitas a prosperarem e se tornarem uma ameaça a seus hospedeiros e a nós mesmos.
Um destes casos é o camarada de hoje, Puccinia graminis, um fungo que causa uma doença em cereais chamada de ferrugem-do-colmo. Conhecida desde a antiguidade, já que já foi mencionada em trabalhos como os de Aristóteles, esta espécie se tornou um problema sério em séculos mais recentes após a expansão da agricultura.
A ferrugem vermelha aparecendo em plantas de trigo são um sinal de infecção.
A ferrugem-do-colmo pertence à ordem Pucciniales, um grupo de fungos conhecidos como ferrugens. Esse nome se refere à aparência que causam nas plantas ao infectá-las, pois partes do tronco e das folhas podem ficar com aparência enferrujada. Apesar de a ferrugem-do-colmo poder infectar várias espécies, seus hospedeiros mais importantes, ao menos de uma perspectiva humana, são o trigo e a cevada, especialmente o trigo.
Quando a ferrugem-do-colmo infecta um pé de trigo, o que ocorre no verão, ela começa a crescer como um micélio dentro do tecido da planta e, após cerca de 1 a 2 semanas, começa a produzir pústulas vermelhas como ferrugem que aparecem principalmente na bainha das folhas, apesar de poderem aparecer em qualquer lugar da planta. Essas pútulas são uredínios e contêm muitos esporos pedunculados chamados de urediniósporos.
Seção transversal de uma folha de trigo mostrando os urediniósporos pedunculados dentro de um uredínio. Foto de Jon Houseman.*
Os urediniósporos são células dicarióticas, isto é, eles têm dois núcleos haploides dentro deles, e são facilmente disseminados pelo vento. Quando caem num novo pé de trigo, germinam para produzir um novo micélio e infectam o novo hospedeiro. Dessa forma o fungo pode se espalhar assexuadamente por uma grande área. Pés de trigo infectados são frequentemente menores, produzem menos grãos ou grãos menores, e às vezes podem até morrer se a infecção for severa demais.
Ao final do ciclo de vida do trigo, o fungo produz outras estruturas, chamadas télios, que produzem uma forma diferente de esporos dicarióticos conhecidos como teliósporos. Os télios possuem uma cor preta e a doença é também conhecida como ferrugem preta por causa disso. Os teliósporos podem sobreviver ao inverno sem um hospedeiro. Durante esse período, seus dois núcleos se fundem e, quando a primavera chega, o teliósporo passa por meiose e produz quatro esporos conhecidos como basidiósporos.
Os télios pretos que produzem os teliósporos têm uma aparência bem diferente dos uredínios sob o microscópio. Foto de John Houseman.*
Os basidióspros são levados pelo vento até atingirem o chamado hospedeiro alternado, uma planta na qual o fungo se reproduz sexuadamente. Este hospedeiro é frequentemente um arbusto do gênero Berberis (bérberis). Quando o basidiósporo cai sobre uma folha de bérberis, germina e produz um micélio haploide. Esse micélio origina estruturas conhecidas como pícnios que produzem tanto hifas femininas conhecidas como hifas receptivas quanto esporos masculinos conhecidos como picniósporos. Os picniósporos são cobertos por um néctar pegajoso que atrai insetos. À medida que esses insetos se movem de uma planta para outra explorando esse presente açucarado, carregam consigo os picniósporos.
Seção transversal de uma folha de bérberis mostrando os pícnios na superfície superior e os grandes écios na superfície inferior. Foto de Jon Houseman.*
Quando um picniósporo encontra uma hifa receptiva de outro fungo, os dois se fundem e crescem num micélio dicariótico que produzirá novamente uma estrutura diferente, o écio, que contém novamente outro tipo de esporo, o eciósporo. Os eciósporos são levados pelo vendo das folhas de bérberis para as plantas de trigo, onde germinam e recomeçam o ciclo.
Écios na superfície inferior da folha de bérberis. Foto de Marina Gorbunova.**
Esse é um ciclo complexo e incrível, né? Contudo humanos odeiam a ferrugem-do-colmo desde tempos antigos. Seus efeitos no trigo são tão importantes que os romanos até tinham um festival inteiro, a Robigália, dedicado a preveni-la. Era um festival horrendo que incluía o sacrifício de um pobre e inocente cão a Robigus, o “deus da ferrugem”.
Na história recente, esse fungo causou sérios danos a plantações de trigo pelo mundo todo diversas vezes, especialmente na Europa, na Ásia e na África. Muitos cultivares modernos foram selecionados para serem resistentes à ferrugem-do-colmo, mas o fungo também evolui, frequentemente originando novas linhagens que são capazes de atravessar a resistência adquirida pelas plantas.
Mas se o fungo precisa infectar plantas de bérberis para completar seu ciclo, não seria só questão de evitar ter bérberis perto de plantações de trigo? Mais ou menos. Mas lembre-se de que quando o fungo está infectando um pé de trigo, ele produz urediniósporos, os quais podem infectar novos pés de trigo. Se você plantar diferentes variedades de trigo que crescem em diferentes épocas do ano, o fungo pode seguir infectando novas plantas assexuadamente sem precisar de qualquer bérberis.
Nós, humanos, somos mesmo bons em tornar as doenças mais virulentas ao tentar combatê-las. Não somos?
Lewis, C. M., Persoons, A., Bebber, D. P., Kigathi, R. N., Maintz, J., Findlay, K., … & Saunders, D. G. (2018). Potential for re-emergence of wheat stem rust in the United Kingdom. Communications biology, 1(1), 1-9. https://doi.org/10.1038/s42003-018-0013-y
Nos anos 1930, uma doença devastadora se espalhou pelas populações atlânticas da fita-do-mar-comum, matando 90% das plantas. A doença começou destruindo os cloroplastos das plantas, tornando as folhas brancas, e em seguida causando apodrecimento dos tecidos que se espalhou até a planta inteira morrer.
Apesar de o agente causador da doença já ter sido sugerido naquela época, ele só foi confirmado durante uma recorrência menor nos anos 1980. O responsável é um parasita marinho “fungoide” conhecido como Labyrinthula zosterae, o qual eu decidi chamar de labiríntula-da-fita-do-mar.
Marcas escuras de tecido apodrecido na fita-do-mar-comum causadas pela labiríntula-da-fita-do-mar. Extraído de Short (2014).
A labiríntula-da-fita-do-mar pertence a um grupo peculiar de organismos que foram inicialmente classificados como um grupo estranho de bolores-limosos (mixomicetos), mas que agora se sabe que são heterocontos, portanto mais proximamente relacionados a diatomáceas, algas marrons e oomicetos, por exemplo. Eles são organismos coloniais e sua colônia é bem interessante em seu arranjo. As células vegetativas individuais são em forma de fuso e medem de 15 a 20 µm de comprimento por 3 a 5 µm de largura. Elas frequentemente são translúcidas, às vezes amarelo-claras, e são preenchidas com gotículas de lipídios.
As células produzem juntas uma rede formada por ectoplasma (citoplasma) que é excretado das células e é rodeado por uma membrana celular, então se tornando tipo uma grande célula em forma de labirinto dentro da qual as células individuais vivem. Esta rede não tem parede celular nem organelas, no entanto. As células podem deslizar através dessa rede, usando-a como uma “estrada”.
Células vegetativas da labiríntula-da-fita-do-mar. A rede ectoplasmática pode ser vista como finas extensões brancas. Extraído de theredshrimp.com.
Há duas formas de reprodução conhecidas até agora. A primeira é simplesmente por divisão celular através de mitose. Outra forma de reprodução é por zoosporulação, que começa com várias células vegetativas se agregando e fundindo em uma estrutura plasmodial única. Mais tarde, este plasmódio se divide de volta em grandes presporângios redondos que então se dividem internamente em oito zoósporos, que são então liberados no ambiente. Os zoósporos são células com dois flagelos e um estigma (ocelo), bem similares às células típicas de outros heterocontos, e vão se diferenciar de volta em células vegetativas.
A infecção da fita-do-mar-comum pela labiríntula-da-fita-do-mar ocorre por contato direto de uma planta saudável com uma infectada ou com partes soltas infectadas. O parasita age dissolvendo a parede celular e se espalhando pelos tecidos, destruindo as células e provavelmente se alimentando delas. Estudos recentes, no entanto, descobriram que a labiríntula-da-fita-do-mar é muito comum em populações de fitas-do-mar, mas a maioria das linhagems não é muito virulenta. De fato, é provável que a labiríntula-da-fita-do-mar seja parte da biota protista “normal” associada à fita-do-mar-comum e só se torna uma ameaça quando a planta passa por algum tipo de estresse induzido por outras condições.
De qualquer forma, ainda há muito a ser descoberto sobre esse protista e suas intenções duvidosas.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Brakel, J., Werner, F. J., Tams, V., Reusch, T. B., & Bockelmann, A. C. (2014). Current European Labyrinthula zosterae are not virulent and modulate seagrass (Zostera marina) defense gene expression. PLoS One, 9(4), e92448. https://doi.org/10.1371/journal.pone.0092448
Muehlstein, L. K., Porter, D., & Short, F. T. (1991). Labyrinthula zosterae sp. nov., the causative agent of wasting disease of eelgrass, Zostera marina. Mycologia, 83(2), 180-191. https://doi.org/10.1080/00275514.1991.12025994
As bactérias estão entre os organismos mais diversos na Terra e desempanham todos os tipos de papéis ecológicos, apesar de elas serem mais frequentemente associadas a doenças pelo ser humano médio. Isso é, claro, devido ao fato de a maioria das bactérias que tem influência direta e perceptível na vida humana ser, de fato, patogênica, frequentemente parasita. Hoje vou apresentar uma destas bactérias patogênicas, mas não uma que infecta humanos.
Atualmente conhecida como Flavobacterium psychrophilum, esta espécie é conhecida por infectar peixes de água doce causando uma doença conhecida como “doença da água fria” (DAF). Como resultado, vou chamar a espécie de bactéria-da-doença-da-água-fria, ou bactéria-da-DAF, de forma abreviada.
O aspecto típico das bactérias-da-DAF vistas sob o microscópio. Extraído de Cipriano & Holt (2005).
A bactéria-da-DAF é uma bactéria com o típico formato de bastão, medindo até 1 µm de largura e até 5 µm de comprimento. Ela não possui nenhum tipo de flagelo ou outra estrutura que a ajude a se mover, mas pode se mover por deslizamento, apesar de este movimento ser incrivelmente lento e muito difícil de ser observado. Quando cultivada em meio de cultura, ela produz pequenas colônias amarelas de 3 mm de diâmetro com margens finas.
Várias colônias de bactérias-da-DAF em meio de cultura. Créditos a Eva Säker (SVA) & Karl-Erik Johansson (BVF, SLU & SVA).*
Vivendo em água doce, a bactéria-da-DAF prefere águas frias com temperaturas de 16°C ou menos, com a temperatura ótima sendo 13°C. Elas crescem em todo tipo de tecido do corpo de peixes, como a pele, brânquias, muco e órgãos internos como cérebro, rins, baço e órgãos sexuais. Seus hospedeiros preferidos são peixes salmonoides, como os salmões e trutas, mas ela também pode eventualmente ser encontrada em outros peixes.
A bactéria-da-DAF é uma bactéria aeróbica, mas não é capaz de usar carboidratos como fonte de carbono, se alimentando, em vez disso, de peptídeos. Assim, ela secreta enzimas nos tecidos do hospedeiro que degradam suas proteínas para formar peptídeos, causando dano estrutural.
Peixes infectados apresentam erosão tecidual, que geralmente começa na nadadeira caudal e eventualmente se espalha. As nadadeiras ficam escuras, rasgadas, partidas ou em frangalhos e podem ser completamente perdidas. Ulcerações aparecem na pele, especialmente em volta da mandíbula, e os peixes apresentam problemas comportamentais como nado em espiral e letargia. A infecção geralmente mata o peixe, mas às vezes uma infecção mais fraca e crônica pode acontecer, a qual, no entanto, ainda causa consideráveis mudanças comportamentais no hospedeiro.
Lesões causadas por Flavobacterium psychrophilum na truta-arco-íris Oncorhynchus mykis (A) e no salão-coho Oncorhynchus kisutsch. Extraído de Starliper et al. (2011).
As bactérias são frequentemente transmitidas de peixe para peixe pelo contato direto de um com o outro, mas peixes adultos infectados também podem passar a infecção diretamente para a prole através de ovos infectados. A infecção pode ser tratada nos estágios iniciais usando o antibiótico oxitetraclina ou pela adição de cátions quaternários de amônio na água.
Em ambientes naturais, os problemas causados por esta infecção raramente são problemáticos e o dano é mais frequentemente visto em fazendas de peixes, onde os pobres peixes são forçados a viver em alta densidade, o que aumenta o sucesso da bactéria. Aparentemente nativa da América do Norte, onde foi descoberta dos anos 1940, a bactéria foi espalhada via piscicultura pelo mundo todo nas décadas seguintes. Nós humanos, portanto, somos mais uma vez a principal razão de esta espécie ter se tornado um problema em escada global.
– – –
Referências:
Cipriano, R. C., & Holt, R. A. (2005). Flavobacterium psychrophilum, cause of bacterial cold-water disease and rainbow trout fry syndrome. Kearneysville, WV: US Department of the Interior, US Geological Survey, National Fish Health Research Laboratory.
Langevin, C., Blanco, M., Martin, S. A., Jouneau, L., Bernardet, J. F., Houel, A., … & Boudinot, P. (2012). Transcriptional responses of resistant and susceptible fish clones to the bacterial pathogen Flavobacterium psychrophilum. PLoS One, 7(6), e39126. https://doi.org/10.1371/journal.pone.0039126
Starliper, C. E. (2011). Bacterial coldwater disease of fishes caused by Flavobacterium psychrophilum. Journal of Advanced Research, 2(2), 97-108. https://doi.org/10.1016/j.jare.2010.04.001
Semana passada apresentei a adorável bolacha-da-praia-de-cinco-furos, um equinodermo muito comum ao longo da Costa Atlântica das Américas. Mas como todos sabemos, as espécies raramente vivem sozinhas. Todo tipo de associação existe entre organismos, e nossa espécie de hoje é uma que vive proximamente associada com a bolacha-da-praia-de-cinco-furos, o caranguejinho-da-bolacha-da-praia, Dissodactylus mellitae.
O caranguejinho-da-bolacha-da-praia é, como o nome sugere, um caranguejo. Um caranguejo bem bem pequeno de fato. Machos adultos atingem até 3.5 mm de tamanho e as fêmeas não ficam maiores que 4.5 mm. Eles possuem uma cor amarelo-clara a branca, às vezes com um padrão de marcas escuras complexas no dorso.
O minúsculo caranguejinho-da-bolacha-da-praia. Créditos ao Naturalis Biodiversity Center.*
O habitat natural do caranguejinho-da-bolacha-da-praia é a superfície de bolachas-da-praia, especialmente a bolacha-da-praia-de-cinco-furos. Como eles são muito pequenos, vivem confortavelmente entre os pelos e espinhos do hospedeiro, mais comumente no lado ventral, protegidos da luz e de possíveis predadores.
Por algum tempo não se sabia se a relação entre as duas espécies era de comensalismo, onde o caranguejo apenas come com a bolacha-da-praia, ou de parasitismo, onde o caranguejo rouba alimento da bolacha-da-praia ou se alimenta da bolacha-da-praia em si. Análises do conteúdo estomacal dos caranguejos revelaram que até 80% da sua dieta consiste de tecidos da bolacha-da-praia, de forma que a relação provavelmente é de parasitismo. De fato, foi demonstrado que a presença dos caranguejos reduz o número de ovos que as bolachas-da-praia fêmeas produzem.
Caranguejinho-da-bolacha-da-praia no seu hospedeiro. Créditos ao Naturalis Biodiversity Center.*
O número máximo de caranguejos observados em uma só bolacha-da-praia foi 10, mas esse número depende muito do estágio de vida do caranguejo. No verão, os jovens frequentemente preferem viver juntos, compartilhando o mesmo hospedeiro, mas à medida que crescem se disperam e preferem uma vida solitária, sem compartilhar o hospedeiro com outros da mesma espécie. Quando estão sexualmente maduros, eles frequentemente compartilham o hospedeiro com outro caranguejo do sexo oposto, assim facilitando a reprodução. Contudo os machos parecem ser muito mais comuns da população, então machos vivendo com outros machos são mais comuns que machos vivendo com fêmeas.
A reprodução parece ocorrer no fim do verão e outono ao longo da costa da América do Norte, depois do qual o número de caranguejos adultos começa a diminuir. Os jovens começam a aparecer no fim da primavera, buscando avidamente bolachas-da-praia para colonizar.
No mar, diferentes espécies se associam ainda mais frequentemente que na terra. E nós sabemos que onde quer que haja vida, haverá outra vida para parasitá-la.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
George, S. B., & Boone, S. (2003). The ectosymbiont crab Dissodactylus mellitae–sand dollar Mellita isometra relationship. Journal of Experimental Marine Biology and Ecology, 294(2), 235-255. https://doi.org/10.1016/S0022-0981(03)00271-5
Semana passada aprendemos sobre a formiga-lava-pés-vermelha e o incômodo que ela é na América do Norte, onde foi introduzida quase um século atrás e se tornou uma ameaça para ecossistemas e pessoas. Como pesticidas químicos costumam ser prejudicais para espécie que não são a alvo, alternativas mais espécie-específicas são frequentemente procuradas. Uma dessas é controle biológico, o qual também pode ser problemático se usado incorretamente.
A espécie de hoje é um desses agentes biológicos e pode ajudar a controlar a invasão da formiga-lava-pés-vermelha. Chamada Kneallhazia solenopsae, que eu decidi apelidar de microsporídio-interno-das-lava-pés, esta espécie pertence à divisão Microsporidia, que são fungos unicelulares. Eu já apresentei um deles aqui muitos anos atrás, a glúgea-de-solha.
O microsporídio-interno-das-lava-pés é conhecido por infectar formigas-lava-pés, incluindo a formiga-lava-pés-vermelha. Ele tem um ciclo de vida bem complexo com vários estágios e que não é completamente entendido ainda.
A infecção mais séria ocorre nas rainhas, nas quais o estágio de vida conhecido como esporo dicariótico do tipo 2 (DK do tipo 2) infecta seu tecido adiposo. Este é um estágio do microsporídio no qual a célula tem dois núcleos, daí o nome. Outras formigas adultas também são infectadas de forma similar. A infecção se espalha lentamente, causando hipertrofia do tecido adiposo e, em estágios avançados, faz esse tecido ser completamente substituído por massas de esporos, impedindo o desenvolvimento e a reprodução da rainha. Contudo demora muito tempo para o microsporídio causar uma doença tão grave. Isso não significa que ele não está fazendo nada para piorar as coisas antes disso, no entanto.
Enquanto o tecido adiposo da rainha está sendo lentamente consumido pelos esporos DK do tipo 2, muitos deles se convertem em outro estágio, o octósporo, assim chamado porque ocorre numa vesícula que contem oito deles. Essas vesículas são relativamente grandes se comparadas a outros estágios e são consideradas o estágio infeccioso. Uma formiga morta ou machucada, que é frequentemente canibalizada por outras, libera as vesículas no ambiente. Elas precisam ser ingeridas para que a infecção ocorra, mas somente as larvas de quarto ínstar são capazes de engoli-las, porque todos os outros estágios larvais, bem como os adultos, possuem um “filtro” na boca que os previne de ingerir partículas maiores que 0.8 µm e as vesículas são maiores que isso.
Várias vesículas de octósporos e um esporo DK do tipo 2 no microsporídio-interno-das-lava-pés. Foto de David Williams.*
Dentro das larvas de quarto ínstar, as vesículas liberam os octósporos que infectam a larva e se fundem, transformando-se em outro tipo de célula com dois núcleos, os esporos DK do tipo 1. Esses esporos só são encontrados em larvas de quarto ínstar e pupas e se supõe que se transformem em esporos DK do tipo 2 durante o desenvolvimento para a forma adulta, onde podem produzir novos octósporos.
Mas essa não é a única forma pela qual o microsporídio se espalha pela colônia. Voltando às rainhas, os esporos DK do tipo 2 também infectam os ovários e, depois que a rainha é inseminada, se trasnformam em outro estágio, o chamado megásporo. Os megásporos infectam os ovos e são incorporados no embrião e de lá passam diretamente para larvas de primeiro ínstar, onde se desenvolvem em esporos DK do tipo 1 a medida em que crescem.
Então a única forma de uma formiga se infectar é se ela recebe o parasita diretamente da rainha no ovo ou se ingere vesículas de octósporos como larva de quarto ínstar? Não. E a razão está no fato de que as formigas realizam trofalaxia, que é a transferência de alimento de um indivíduo para outro, isto é, as formigas vomitam uma na boca da outra. Assim, se um adulto troca alimento com uma larva de quarto ínstar que liberou os octósporos no tubo digestivo, ele pode se infectar também.
É um ciclo bem complicado, certo? Mas ele parece ser bem afinado para que o microsporídio seja um parasita bem-sucedido das formigas-lava-pés. Mas ele é um controle biológico eficiente? Bem, eu diria que ele pode ajudar a retardar a disseminação das formigas-lava-pés, mas como seus efeitos de causar esterilidade levam muito tempo para surgir, ele não causará a remoção das formigas em áreas que já foram invadidas.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Knell, J. D., Allen, G. E., & Hazard, E. I. (1977). Light and electron microscope study of Thelohania solenopsae n. sp.(Microsporida: Protozoa) in the red imported fire ant, Solenopsis invicta. Journal of Invertebrate Pathology, 29(2), 192-200. https://doi.org/10.1016/0022-2011(77)90193-8
Oi, D. H., & Williams, D. F. (2002). Impact of Thelohania solenopsae (Microsporidia: Thelohaniidae) on polygyne colonies of red imported fire ants (Hymenoptera: Formicidae). Journal of Economic Entomology, 95(3), 558-562. https://doi.org/10.1603/0022-0493-95.3.558



 Esta obra está licenciada sob uma Licença Creative Commons de Atribuição Não Comercial 4.0 Internacional
Esta obra está licenciada sob uma Licença Creative Commons de Atribuição Não Comercial 4.0 Internacional