por Piter Kehoma Boll
Sete anos atrás eu discuti a posição filogenética de Acoelomorpha e seus parentes próximos, Xenoturbella, que juntos formam o clado Xenacoelomorpha. Sendo animais bilaterais muito simples que não possuem quase nenhuma das principais estruturas comuns à maioria dos outros bilaterais, sua posição filogenética exata é geralmente considerada basal dentro de Bilateria, mas a ideia de que são deuterostômios foi levantada depois que alguns estudos moleculares os agruparam dentro do clado Ambulacraria, que inclui equinodermos e hemicordados.
Pertencer aos deuterostômios significaria que Xenacoelomorpha sofreu uma gigantesca simplificação de sua anatomia. Lá em 2013, quando escrevi o outro artigo, isso estava causando bastante controvérsia, mas, pouco depois, novos estudos moleculares confirmaram a posição basal de Xenacoelomorpha e ficou meio que bem aceito que eles eram, de fato, o clado mais basal de Bilateria.

Mas uma vez problemático, sempre problemático.
Em 2019, um novo estudo que tentou antecipar os erros sistemáticos durante estudos de filogenia molecular, tal como atração de ramos longos, concluiu que a posição basal de Xenacoelomorpha é um artefato e que, quando se tenta minimizar os erros, sua posição como grupo irmão de Ambulacraria se torna clara. Contudo a árvore deles também sugere que Deuterostomia não seja monofilético, já que os cordados aparecem como grupo irmão de Protostomia e Xenacoelomorpha+Ambulacraria é o grupo mais basal, isto é, o grupo irmão do resto dos Bilateria. Contudo a ideia de Deuterostomia não ser monofilético é bem inesperada.
Como mencionei no meu último post, o principal problema em Xenacoelomorpha aparecer dentro de Deuterostomia tem a ver com sua supersimplificação. Eles não têm quase nada que um bilateral típico tem. O que os teria forçado a se tornarem tão simples?
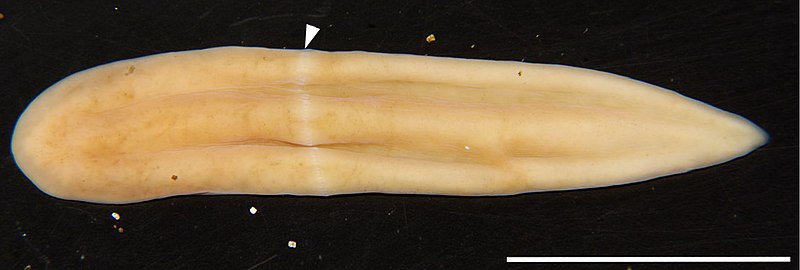
Outro estudo recente sugere que, no caso de Xenoturbella ao menos, isso pode ser o resultado de seus hábitos de se enterrar num substrato macio. Eles comparam Xenoturbella a nudibrânquios, entre os quais algumas espécies possuem estilos de vida similares. Um desses nudibrânquios, Xenocratena, foi na verdade descoberto mais ou menos na mesma época que Xenoturbella vivendo no mesmo ambiente que ela. Eles possuem uma anatomia pedomórfica (mais simplificadas, “de bebê”) em comparação a outros nudibrânquios. Contudo ela não é de forma alguma tão simples quanto a de Xenoturbella.

Por outro lado, há outro gênero de nudibrânquios que é de fato supersimplificado, Pseudovermis, e ele vive enterrado em substrato macio também. Análises moleculares revelaram que Pseudovermis não é proximamente relacionado a Xenocratena, mas a Cumanotus, outro nudibrânquio que se enterra, o que sugere que esta simplificação ocorreu duas vezes entre os nudibrânquios.
Isso não é evidência de que Xenoturbella é um deuterostômio simplificado, mas e um bom argumento. Mas e quanto às simplificações em Acoelomorpha? Penso que se Xenoturbella não fosse proximamente relacionada a Acoelomorpha, eu estaria mais disposto a aceitar essa hipótese. Meu coração tende para a hipótese de Xenacoelomorpha basal, no entanto. Contudo, como qualquer cientista, aceitarei Xenacoelomorpha como deuterostômios se forem apresentadas evidências suficientes.
Xenoturbella é sempre o problema principal nesta equação. O sistema nervoso de Acoelomorpha, por exemplo, apesar de simplificado, tem meio que o padrão básico encontrado em todos os bilaterais e poderia ter evoluído do anel oral de um ancestral similar a um cnidário de acordo com algumas hipóteses. En Xenoturbella, no entanto, o sistema nervoso é muito mais estranho, sendo formando por uma rede difusa simples de neurônios abaixo da pele. Acho que abordar a organização do sistema nervoso em todos esses grupos é um bom tópico para outra postagem.
Se há uma coisa, na minha opinião, que faz a posição de Xenacoelomorpha dentro Deuterostomia ser algo convincente é o fato de muitas características de Deuterostomia parecerem ser mais primitivas dentro de Bilateria quando comparadas àquelas de Protostomia, de forma que a posição de Xenacoelomorpha entre os Deuterostomia é mais plausível que sua posição dentro de Protostomia (apesar de isso nem mesmo ser mais considerado uma possibilidade). Tendemos a pensar que deuterostômios são mais “derivados” simplesmente porque humanos são deuterostômios. Mas esta discussão também e assunto para outra postagem.
O que você acha? Você é time basal ou time deuterostômio?
– – –
Você também pode gostar:
Peixes-Bruxas: outra dor de cabeça filogenética
Xenoturbella: um grupo crescente de esquisitões
– – –
Curta nossa página no Facebook!
Siga-me (@piterkeo) no Twitter!
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com
Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Cannon JT, Vellutini BC, Smith J, Ronquist F, Jonfelius U, Hejnol A (2016) Xenacoelomorpha is the sister group to Nephrozoa. Nature 530: 89–93. doi: 10.1038/nature16520
Jondelius U, Raikova OI, Martinez P (2019) Xenacoelomorpha, a Key Group to Understand Bilaterian Evolution: Morphological and Molecular Perspectives. In: Pontarotti P (ed) Evolution, Origin of Life, Concepts and Methods. Cham: Springer International Publishing, . pp. 287–315. doi: 10.1007/978-3-030-30363-1_14
Martynov A, Lundin K, Picton B, Fletcher K, Malmberg K, Korshunova T (2020) Multiple paedomorphic lineages of soft-substrate burrowing invertebrates: parallels in the origin of Xenocratena and Xenoturbella. PLoS ONE 15(1): e0227173. doi: 10.1371/journal.pone.0227173
Philippe H, Poustka AJ, Chiodin M, et al. (2019) Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria. Current Biology 29(11):1818–1826. doi: 10.1016/j.cub.2019.04.009
– – –
* Esta obra está licenciada sob uma Licença Creative Commons de Atribuição 4.0 Internacional.
Esta obra está licenciada sob uma Licença Creative Commons de Atribuição 4.0 Internacional.