As pessoas amam nomear criaturas marinhas fazendo analogias com coisas encontradas em terra. A espécie de hoje é mais uma desse tipo, sendo a mais bem conhecida espécie das chamadas borboletas-do-mar e, portanto, conhecida como a borboleta-do-mar-comum. Ela não tem nada a ver com borboletas, no entanto, e seu nome científico, Limacina helicina, a descreve melhor.
A borboleta-do-mar-comum é um molusco, mais precisamente um gastrópode e, como tem concha, é um “caramujo”. Ela não se arrasta pelo fundo como a maioria dos caramujos, no entanto. Com uma concha espiral medindo somente 10 mm de diâmetro em média, ela vive na coluna d’água e às vezes é descrita como uma espécie planctônica. Ela pode nadar por conta própria, no entanto, porque seu pé carnoso é transformado em duas expansões chamadas parapódios que agem como duas grandes nadadeiras. Sua concha é transparente e as partes moles são principalmente roxas, apesar de os parapódios serem quase transparentes também.
A borboleta-do-mar-comum é mesmo uma criatura linda, não é? Foto de Russ Hopcroft, University of Alaska, Fairbanks.
O habitat da borboleta-do-mar-comum inclui as águas frias da região ártica, incluindo o Oceano Ártico e áreas vizinhas dos oceanos Pacífico e Atlântico. No Pacífico, ela pode ocorrer para o sul até o Japão e regiões mais ao norte dos Estados Unidos. Espécimes maiores tendem a habitar águas mais profundas, até 150 m de profundidade, enquanto os menores vivem mais perto da superfície, até uma profundidade de 50 m. Até bem recentemente, pensava-se que a borboleta-do-mar-comum também habitasse águas antárticas, mas estudos moleculares revelaram que as populações em torno da Antártica pertencem a outra espécie, Limacina antarctica.
Veja como elas conseguem bater suas asas com velocidade,
A dieta da borboleta-do-mar-comum inclui várias criaturas planctônicas menores, especialmente crustáceos pequenos, como náuplios (larvas) de copépodes, bem como dinoflagelados, ciliados e diatomáceas. Juvenis da própria espécie também são comuns, às vezes compondo o segundo item mais comum da dieta. Para capturar a comida, elas produzem uma teia esférica de muco que flutua acima delas na água. Esta teia captura outros organismos na coluna d’água e depois é sugada e comida pela borboleta-do-mar junto com as criaturas capturadas. Esta teia é muito difícil de ser observada durante o dia por causa da refração difusa, mas aparece claramente à noite. Quando perturbada por luz, no entanto, a borboleta-do-mar-comum tende a engolir sua teia rapidamente e afundar para escapar do perigo.
Uma borboleta-do-mar-comum com sua teia de muco esférica vista como uma concentração coval de partículas mais finas logo acima dela. Extraído de Gilmer & Harbinson (1986).
A fina concha da borboleta-do-mar-comum é constituída de aragonita, a qual é altamente solúvel e sensível a mudanças na temperatura e na acidificação da água. Estudos demonstraram que o esperado aumento da acidificação dos oceanos pelas mudanças climáticas induzidas por humanos provavelmente terá um impacto negativo em populações da borboleta-do-mar-comum e espécies relacionadas. Isso é particularmente preocupante em relação à borboleta-do-mar-comum porque ela é uma espécie-chave nos ecossistemas árticos, sendo uma fonte de alimento importante para muitos animais marinhos, como peixes, baleias, aves e até outros moluscos.
Esse caramujinho não vai se dar por vencido tão facilmente, no entanto. Estudos mostraram que o periostraco (a camada externa orgânica da concha) pode impedir que a aragonita se dissolva e que um trauma físico que quebre o periostraco, permitindo contato direto da aragonita com a água, é necessário para causar a dissolução. E mesmo quando isso acontece, a borboleta-do-mar-comum pode compensar construindo novas camadas de aragonita na superfície interna da concha e consegue extrair aragonita da água para esse propósito mesmo quando os níveis na água estão muito baixos.
A borboleta-do-mar-comum é pequena mas também é durona.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Corneau S, Alliouane S, Gattuso JP (2012) Effects of ocean acidification on overwintering juvenile Arctic pteropods Limacina helicina. Marine Ecology Progress Series 456:279–284. doi: 10.3354/meps09696
Comeau S, Jeffree R, Teyssié JL, Gattuso JP (2010) Response of the Arctic pteropod Limacina helicina to projected future environmental conditions. PLoS One 5(6):e11362. doi: 10.1371/journal.pone.0011362
Gilmer RW, Harbinson GR (1986) Morphology and field behavior of pteropod molluscs: feeding methods in the families Cavoliniidae, Limacinidae and Peraclididae (Gastropoda: Thecosomata). Marine Biology 91:47–57. doi: 10.1007/BF00397570
Lischka S, Büdenbender J, Boxhammer T, Riebesell U (2011) Impact of ocean acidification and elevated temperatures on early juveniles of the polar shelled pteropod Limacina helicina: mortality, shell degradation, and shell growth. Biogeosciences 8:919–932. doi: 10.5194/bg-8-919-2011
Sete anos atrás eu discuti a posição filogenética de Acoelomorpha e seus parentes próximos, Xenoturbella, que juntos formam o clado Xenacoelomorpha. Sendo animais bilaterais muito simples que não possuem quase nenhuma das principais estruturas comuns à maioria dos outros bilaterais, sua posição filogenética exata é geralmente considerada basal dentro de Bilateria, mas a ideia de que são deuterostômios foi levantada depois que alguns estudos moleculares os agruparam dentro do clado Ambulacraria, que inclui equinodermos e hemicordados.
Pertencer aos deuterostômios significaria que Xenacoelomorpha sofreu uma gigantesca simplificação de sua anatomia. Lá em 2013, quando escrevi o outro artigo, isso estava causando bastante controvérsia, mas, pouco depois, novos estudos moleculares confirmaram a posição basal de Xenacoelomorpha e ficou meio que bem aceito que eles eram, de fato, o clado mais basal de Bilateria.
Uma versão simplificada da árvore da vida animal mostrando a posição incerta de Xenacoelomorpha. A posição de Placozoa e Ctenophora também não é muito clara.*
Mas uma vez problemático, sempre problemático.
Em 2019, um novo estudo que tentou antecipar os erros sistemáticos durante estudos de filogenia molecular, tal como atração de ramos longos, concluiu que a posição basal de Xenacoelomorpha é um artefato e que, quando se tenta minimizar os erros, sua posição como grupo irmão de Ambulacraria se torna clara. Contudo a árvore deles também sugere que Deuterostomia não seja monofilético, já que os cordados aparecem como grupo irmão de Protostomia e Xenacoelomorpha+Ambulacraria é o grupo mais basal, isto é, o grupo irmão do resto dos Bilateria. Contudo a ideia de Deuterostomia não ser monofilético é bem inesperada.
Como mencionei no meu último post, o principal problema em Xenacoelomorpha aparecer dentro de Deuterostomia tem a ver com sua supersimplificação. Eles não têm quase nada que um bilateral típico tem. O que os teria forçado a se tornarem tão simples?
Outro estudo recente sugere que, no caso de Xenoturbella ao menos, isso pode ser o resultado de seus hábitos de se enterrar num substrato macio. Eles comparam Xenoturbella a nudibrânquios, entre os quais algumas espécies possuem estilos de vida similares. Um desses nudibrânquios, Xenocratena, foi na verdade descoberto mais ou menos na mesma época que Xenoturbella vivendo no mesmo ambiente que ela. Eles possuem uma anatomia pedomórfica (mais simplificadas, “de bebê”) em comparação a outros nudibrânquios. Contudo ela não é de forma alguma tão simples quanto a de Xenoturbella.
Por outro lado, há outro gênero de nudibrânquios que é de fato supersimplificado, Pseudovermis, e ele vive enterrado em substrato macio também. Análises moleculares revelaram que Pseudovermis não é proximamente relacionado a Xenocratena, mas a Cumanotus, outro nudibrânquio que se enterra, o que sugere que esta simplificação ocorreu duas vezes entre os nudibrânquios.
Relações filogenéticas entre nudibrânqios. Veja Pseudovermis and Cumanotus a 2 horas e Xenocratena a 7 horas. Créditos a Martynov et al. (2020).*
Isso não é evidência de que Xenoturbella é um deuterostômio simplificado, mas e um bom argumento. Mas e quanto às simplificações em Acoelomorpha? Penso que se Xenoturbella não fosse proximamente relacionada a Acoelomorpha, eu estaria mais disposto a aceitar essa hipótese. Meu coração tende para a hipótese de Xenacoelomorpha basal, no entanto. Contudo, como qualquer cientista, aceitarei Xenacoelomorpha como deuterostômios se forem apresentadas evidências suficientes.
Xenoturbella é sempre o problema principal nesta equação. O sistema nervoso de Acoelomorpha, por exemplo, apesar de simplificado, tem meio que o padrão básico encontrado em todos os bilaterais e poderia ter evoluído do anel oral de um ancestral similar a um cnidário de acordo com algumas hipóteses. En Xenoturbella, no entanto, o sistema nervoso é muito mais estranho, sendo formando por uma rede difusa simples de neurônios abaixo da pele. Acho que abordar a organização do sistema nervoso em todos esses grupos é um bom tópico para outra postagem.
Se há uma coisa, na minha opinião, que faz a posição de Xenacoelomorpha dentro Deuterostomia ser algo convincente é o fato de muitas características de Deuterostomia parecerem ser mais primitivas dentro de Bilateria quando comparadas àquelas de Protostomia, de forma que a posição de Xenacoelomorpha entre os Deuterostomia é mais plausível que sua posição dentro de Protostomia (apesar de isso nem mesmo ser mais considerado uma possibilidade). Tendemos a pensar que deuterostômios são mais “derivados” simplesmente porque humanos são deuterostômios. Mas esta discussão também e assunto para outra postagem.
O que você acha? Você é time basal ou time deuterostômio?
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Cannon JT, Vellutini BC, Smith J, Ronquist F, Jonfelius U, Hejnol A (2016) Xenacoelomorpha is the sister group to Nephrozoa. Nature 530: 89–93. doi: 10.1038/nature16520
Jondelius U, Raikova OI, Martinez P (2019) Xenacoelomorpha, a Key Group to Understand Bilaterian Evolution: Morphological and Molecular Perspectives. In: Pontarotti P (ed) Evolution, Origin of Life, Concepts and Methods. Cham: Springer International Publishing, . pp. 287–315. doi: 10.1007/978-3-030-30363-1_14
Martynov A, Lundin K, Picton B, Fletcher K, Malmberg K, Korshunova T (2020) Multiple paedomorphic lineages of soft-substrate burrowing invertebrates: parallels in the origin of Xenocratena and Xenoturbella. PLoS ONE 15(1): e0227173. doi: 10.1371/journal.pone.0227173
Philippe H, Poustka AJ, Chiodin M, et al. (2019) Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria. Current Biology 29(11):1818–1826. doi: 10.1016/j.cub.2019.04.009
Fungos apresentam uma variedade de diferentes corpos de frutificação e são quase sempre nomeados com base no que esses corpos de frutificação parecem. Esse é o caso com a espécie de hoje também. Seu nome científico é Aleuria aurantia e ela é comumente conhecida como o fungo-casca-de-laranja. Mas preste atenção: é fungo-casca-de-laranja e não fungo-DA-casca-de-laranja, já que o seguro se referiria a um fungo crescendo em cascas de laranjas, mas não é o caso, já que tal espécie é chamada de bolor-verde.
O fungo-casca-de-laranja é encontrado através da região Holártica, isto é, América do Norte, Europa e Norte da Ásia, e eventualmente em outros lugares, provavelmente por dispersão causada por humanos. Seu habitat principal inclui áreas abertas perto de florestas de coníferas, onde ele vive como saprótrofo, ou seja, como um fungo decompositor com o micélio espalhado através do solo.
Fungo-casca-de-laranja na França. Foto de David Renoult.*
Os corpos de frutificação em forma de copo ou taça aparecem mais frequentemente no outono. Eles possuem uma cor laranja forte causada por carotenoides, especialmente β-caroteno, γ-caroteno e aleuriaxantina. No começo eles são pequenos e bem regulares, mas crescem até 6 cm em diâmetro, quando sua estrutura começa a se tornar irregular ou se rasga, lembrando pedaços de casca de laranja jogados no chão Os esporos que eles produzem ficam sem germinar no solo até a primavera, quando começam a crescer. As temperaturas frias do inverno parecem ser necessárias para os esporos germinarem, já que esporos em culturas de laboratório só germinaram com sucesso após permanecerem congelados por cerca de 3 meses.
O fungo-casca-de-laranja tem sido alvo de estudos farmacológicos devido à presença de uma lectina chamada lectina de Aleuria aurantia (AAL). Lectinas são um grupo de proteínas que se ligam a açúcares ou grupos açúcar de outras moléculas como glicoproteínas e glicolipídios. Elas são encontradas em todo tipo de organismo e possuem funções diferentes. Em animais, por exemplo, elas são importantes para adesão celular. Elas se conectam a glicoproteínas na membrana celular e ligam células vizinhas umas às outras, prevenindo que os tecidos se desfaçam.
Corpos de frutificação achatados e quebrados lembrando casca de laranja nos Estados Unidos. Foto do usuário schwee do iNaturalist.*
Em muitos fungos, bactérias e vírus parasitas, as lectinas são importantes para reconhecer o hospedeiro. Elas também parecem ter algum papel durante a produção de esporos em fungos. A AAL do fungo-casca-de-laranja se liga à fucose, um açúcar que é comumente encontrado na superfície de células de mamífero, insetos e plantas como parte de suas glicoproteínas. O nome fucose vem do fato de este açúcar formar o polissacarídeo fucoidano encontrado em várias espécies de algas-marrons.
A AAL possui vários usos práticos. Devido ao seu comportamento de se ligar à fucose, ela pode ser usada para detectar a presença de fucose em diferentes células. A fucose também está relacionada a alguns tipos de alergia e a AAL vem sendo estudada como um composto para o potencial desenvolvimento de novos antialérgicos.
Mais que suas propriedades farmacológicas, o fungo-casca-de-laranja também é comestível, apesar de não ser tão popular como alimento. Seus corpos de frutificação podem ser comidos frescos ou secados para armazenamento. Devido à presença de carotenoides, o fungo-casca-de-laranja possui potenciais propriedades antioxidantes, apesar de parecer não haver estudos sobre seu valor nutricional.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Fukumori F, Takeuchi N, Hagiwara T, Ohbayashi H, Endo T, Kochibe N, Nagata Y, Kobata A (1990) Primary structure of a fucose-specific lectin obtained from a mushroom, Aleuria aurantia. Journal of Biochemistry 107(2):190–196. doi: 10.1093/oxfordjournals.jbchem.a123024
Ogawa S, Nagao H, Ando A, Nagata Y (2000) Enhancement of ascospore germination from Aleuria aurantia after cold storage. Mycoscience 41:287–289. doi: 10.1007/BF02489686
Ogawa S, Nakajima E, Nagao H, Ohtoshi M, Ando A, Nagata Y (1998) Synthesis of a Lectin in Both Mycelia and Fruit Bodies of the Ascomycete Mushroom Aleuria aurantia. Bioscience, Biotechnology, and Biochemistry 62(5):915–918. doi: 10.1271/bbb.62.915
Roth-Walter F, Schöll I, Untersmayr E, Fuchs R, Boltz-Nitulescu G, Weissenböck A, Scheiner O, Gabor F, Jensen-Jarolim E (2004) M cell targeting with Aleuria aurantia lectin as a novel approach for oral allergen immunotherapy. Journal of Allergy and Clinical Immunology 114(6):1361–1368. doi: 10.1016/j.jaci.2004.08.010
Seaver FJ (1914) North American Species of Aleuria and Aleurina. Mycologia 6(6):273–278. doi: 10.1080/00275514.1914.12020977
Singh U, Bhatt RP, Stephenson SL, Uniyal P, Mehmood T (2017) Wild edible mushrooms from high elevations in the Garhwal Himalaya—II. Current Research in Environmental & Applied Mycology 7(3):208–226. doi: 10.5943/cream/6/2/6
Węgiel J, Kohlmünzer S (2001) Mycelial culture of the fungus Aleuria aurantia and some of its metabolites. Pharmaceutical Biology 39(2):108–112. doi: 10.1076/phbi.39.2.108.6249
Wimmerova M, Mitchell E, Sanchez JF, Gautier C, Imberty A (2003) Crystal structure of fungal lectin six-bladed β-propeller fold and novel fucose recognition mode for Aleuria aurantia Lectin. Journal of Biological Chemistry 278: 27059-27067. doi: 10.1074/jbc.M30264220
Orquídeas são flores ornamentais muito populares e vêm numa grande variedade de cores, tamanhos e formas. Encontrá-las em ambientes naturais não é sempre tão fácil, mas também não é difícil se você estiver visitando uma área de Mata Atlântica no Brasil.
Uma espécie de orquídea que é nativa deste bioma ameaçado é Cattleya coccinea, até muito recentemente conhecida como Sophronitis coccinea, razão pela qual ainda é frequentemente chamada de sofronite, ou Sophronitis, pelos orquidófilos. Mesmo entre orquidófilos, esta espécie geralmente não possui um nome comum, mas acho que “sofronite-vermelha” fica bem, apesar de, bem… quase todas as espécies do antigo gênero Sophronitis serem vermelhas.
Um espécime de Cattleya coccinea crescendo perto dos cânions entre Santa Catarina e o Rio Grande do Sul. Foto de João Gava Just.*
Ocorrendo em áreas de elevação média a alta do Espírito Santo para o sul até o Rio Grande do Sul e áreas vizinhas em Misiones, Argentina, a sofronite-vermelha é uma orquídea relativamente pequena que produz flores solitárias de um vermelho-escarlate brilhante. As duas pétalas são muito mais largas que as três sépalas, apesar de o labelo, a terceira pétala, tubular, ser mais estreita.
Apesar de às vezes mencionada como uma espécie ameaçada, a sofronite-vermelha é a espécie do grupo Sophronitis com a maior população natural e maior distribuição geográfica. Uma espécie proximamente relacionada, a sofronite-da-Mantiqueira, Cattleya mantiqueirae, está numa situação muito mais crítica e foi, inicialmente, considerada uma subespécie da sofronite-vermelha. Análises moleculares recentes, no entanto, questionaram a classificação atual de ambas as espécies e elas podem acabar se tornando novamente uma só espécie ou serem divididas em ainda mais espécies.
A sofronite-vermelha floresce entre o fim do inverno e o começo da primavera, com o pico de floração entre agosto e outubro. A flor, normalmente só uma por vez, não tem cheiro nem néctar. Mesmo assim, a natureza conspícua da flor sugere que a planta seja polinizada por um animal orientado pela visão. Após algumas observações na natureza, o único polinizador identificado foi o beija-flor Chlorostilbon lucidus. A pequena ave visita as flores procurando por néctar, mas, sem achar nada, vai embora bem depressa, em menos de 5 segundos, mas isso é suficiente para levar o pólen de uma flor para outra. Abelhas, que são polinizadores comuns de outras espécies de Cattleya, não parecem ter qualquer interesse nesta espécie.
Apesar da aparência adorável, a sofronite-vermelha é uma mentirosa. Ela atrai um polinizador ingênuo prometendo alguma recompensa, mas faz a pobre criatura ir embora sem nada. Mas beleza nunca foi sinônimo de bondade apesar dos esforços humanos para achar que sim.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências:
Caballero-Villalobos L, Silva-Arias GA, Buzatto CR, Nervo MH, Singer RB (2017) Generalized food-deceptive pollination in four Cattleya (Orchidaceae: Laeliinae) species from Southern Brazil. Flora 234: 195–206. doi: 10.1016/j.flora.2017.07.014
Rodrigues JF, van den Berg C, Abreu AG, Novello M, Veasey EA, Oliveira GCX, Koehler S (2014) Species delimitation of Cattleya coccinea and C. mantiqueirae (Orchidaceae): insights from phylogenetic and population genetics analyses. Plant Systematics and Evolution 301:1345–1359. doi: 10.1007/s00606-014-1156-z
Então, estamos passando por um tipo de apocalipse como todos sabem. Um vírus agressivo e contagioso se espalhou pelo mundo todo e está causando um grande impacto em nossa sociedade, matando milhares de pessoas e acabando com a economia.
Mas não estou aqui para falar sobre como se proteger do vírus e quem é mais vulnerável a ele. Você pode encontrar essa informação virtualmente em todo lugar (mas não acredite nas besteiras que Karen, a anti-vacina, ou o seu tio Airton, o bolsominion, estão espalhando pelo Whatsapp. Isso é pior que o vírus). Igualmente eu não estou aqui para mostrar como essa pandemia é um resultado direto de nossa sociedade capitalista falha e como as porras dos ricos deveriam ser decapitados de uma vez por todas. Não. Eu farei uma abordagem mais biológica e explicarei um pouco sobre o que é um vírus de um ponto de vista estrutural, funcional e taxonômico.
Então vamos começar com o que é um vírus.
Um vírus é basicamente um monte de m… material genético parasita que infecta células para se reproduzir. Os vírus não são bem seres vivos já que não possuem nem células nem metabolismo. No entanto eles precisam de células para se reproduzirem. Todos os vírus são compostos de um pedaço de ácido nucleico (ou DNA ou RNA) e um capsídeo, uma “caixa” que protege o ácido nucleico. O capsídeo geralmente é formando por muitas cópias de uma ou duas proteínas que são codificadas no material genético do vírus. Cada molécula individual de proteína do capsídeo é chamada de capsômero.
Esquema de um vírus helicoidal mostrando o capsídeo em verde e o material genético em azul. Créditos a Anderson Brito.*
O vírus do mosaico do tabaco (Tobacco mosaic virus) que infecta pés de tabaco e outras plantas, possui um capsídeo helicoidal típico.
A maioria dos vírus possui um capsídeo helicoidal ou icosaédrico. Em um capsídeo helicoidal, os capsômeros são dispostos helicoidalmente e formam um tubo alongado e oco dentro do qual o material genético fica localizado. Em capsídeos icosaédricos, os capsômeros se arranjam para formar um icosaedro, isto é, um poliedro com 20 faces, que envolve o material genético.
Esquema de um vírus icosaédrico com um capsídeo icosaédrico (em verde) em volta do material genético (em vermelho). Créditos a Anderson Brito.*
Adenovírus são um exemplo de vírus com capsídeo icosaédrico. Foto de Graham Beards.**
Muitos vírus possuem uma camada adicional, o envelope, que envolve o capsídeo. O envelope é uma camada bilipídica atravessada por glicoproteínas, como as membranas celulares dos organismos vivos, e é formada pela membrana celular ou por uma membrana interna da célula em que o vírus nasceu. Ele é, portanto, muito similar à membrana celular do hospedeiro do vírus.
Esquema de um vírus icosaédrico envelopado. A camada bilipídica é mostrada em cinza e as glicoproteínas em laranja. Créditos a Anderson Brito.*
Considerando o tipo de ácido nucleico encontrado em vírus, eles podem ser classificados em três grupos principais: vírus de DNA, vírus de RNA e retrovírus.
Vírus de DNA possuem DNA como seu ácido nucleico. Quando eles infectam a célula, eles seguem até o núcleo, onde dependem inteiramente da maquinaria da célula para se reproduzirem, ou seja, eles usam a DNA-polimerase do hospedeiro para produzir novas cadeias de DNA e a RNA-polimerase do hospedeiro para construir RNA viral que será, por sua vez, convertido em proteínas do capsídeo usando os ribossomos da célula. Os vírus de DNA sofrem pouca mutação porque as enzimas DNA-polimerase possuem uma habilidade revisora, isto é, elas podem detectar erros durante a replicação e consertá-los. Vírus como Herpesvirus (que causam herpes e catapora), Poxvirus (que inclui o agora extinto vírus da Varíola) e Adenovirus são todos vírus de DNA.
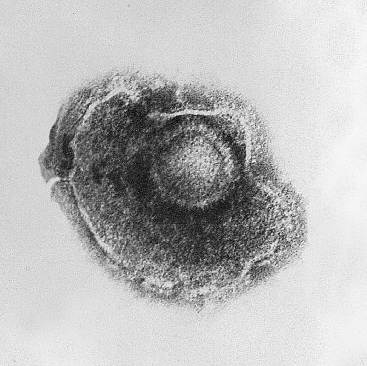
Human alphaherpesvirus 3 (HHV-3) é um vírus de DNA icosaédrico e envelopado que causa catapora e herpes-zóster (cobreiro) em humanos.
Vírus de RNA, também chamados de ribovírus, por outro lado, possuem RNA como seu ácido nucleico. Quando eles infectam uma célula, geralmente permanecem no citoplasma. Diferente dos vírus de DNA, os vírus de RNA possuem sua própria RNA-polimerase e a usam para produzir novas cópias, mas ainda dependem dos ribossomos do hospedeiro para construir os capsídeos e fazer novas cópias da RNA-polimerase. Como enzimas RNA-polimerase não possuem a habilidade revisora das DNA-polimerases, vírus de RNA sofrem mutação rapidamente. Muitas doenças humanas são causadas por vírus de RNA, incluindo resfriado comum, gripe, ebola, febre amarela, dengue, zika, hepatite C, raiva, poliomielite, sarampo, bem como COVID-19, causada pelo atual coronavírus apocalíptico.
Yellow fever virus, o vírus da febre amarela, é um vírus de RNA icosaédrico e envelopado.
Retrovírus, o último tipo de vírus, possuem RNA como seu ácido nucleico. Contudo, diferente dos vírus de RNA, os retrovírus não produzem cópias novas diretamente de seu RNA usando uma RNA-polimerase. Em vez disso, eles possuem outro tipo de enzima, chamada transcriptase reversa, que constrói uma fita de DNA a partir do RNA. Esse DNA viral é então incorporado no DNA da célula hospedeira por uma enzima integrase. Retrovírus, portanto, mudam o genoma do hospedeiro, isto é, eles produzem um “híbrido” deles mesmos com o hospedeiro. A célula infectada então transcreve o DNA viral de volta em RNA, fazendo várias cópias e permitindo que o vírus se reproduza. Os retrovírus mais famosos a infectar humanos são o vírus da imunodeficiência humana (Human immunodeficiency virus, HIV) e o vírus da Hepatite B (Hepatitis B virus).
Human Imunodeficiency Virus 1 (HIV-1) é um retrovírus icosaédrico envelopado que causa AIDS em humanos.
Mas agora vamos nos focar na nossa atual celebridade, SARS-CoV-2, coloquialmente conhecido como o coronavírus. Este vírus, que está causando o apocalipse atual, é uma nova linhagem, descoberta no final de 2019, do coronavírus relacionado à síndrome respiratória aguda severa ((Severe acute respiratory syndrome-related coronavirus (SARSr-CoV)). O surto anterior de SARS entre 2002 e 2004 foi causado por outra linhagem desta mesma espécie, SARS-CoV, agora frequentemente referida como SARS-CoV-1. O vírus pertence ao gênero Betacoronavirus e à família Coronaviridae. Todos os membros da família Coronaviridae são frequentemente chamados de “coronavírus” e as espécies atualmente conhecidas infectam aves e mamíferos.
SARS-COV-2 com cores artificiais mostrando a “coroa” (em laranja) formada pelas glicoproteínas em forma de bastão do envelope.
Coronavírus são vírus de RNA, como mencionei acima, e possuem um capsídeo helicoidal, bem como um envelope. O envelope contém grandes glicoproteínas em forma de bastão que aparecem como projeções na superfície do vírus e, em micrografias eletrônicas, se parecem com a coroa solar, de onde o nome coronavirus (corona, coroa em latim). O envelope é construído a partir da membrana do retículo endoplasmático do hospedeiro e possui glicoproteínas de origem viral, incluindo as glicoproteínas em forma de bastão que caracterizam esses vírus.
A presença deste envelope em torno do capsídeo possui vantagens e desvantagens para os coronavírus e qualquer outro vírus envelopado. Como este envelope é basicamente um pedaço da célula hospedeira, vírus envelopados podem se esgueirar para dentro de hospedeiros novos mais facilmente porque o sistema imunológico leva algum tempo para reconhecê-los como invasores, visto que eles estão “em pele de hospedeiro”. Por outro lado, este envelope é muito frágil quando exposto ao ambiente externo e degrada muito rapidamente, de forma que o vírus precisa de contato próximo entre um hospedeiro infectado e um novo hospedeiro para se espalhar. Também é por isso que lavar as mãos com sabão mata o vírus tão facilmente. Se o vírus não estivesse envelopado, isto é, tivesse somente seu capsídeo, ele seria muito mais resistente.
As glicoproteína em forma de bastão do envelope viral também são responsáveis pela habilidade do vírus de infectar. Estas glicoproteínas se conectam à enzima conversora de angiotensina 2 (angiotensin-converting enzyme 2 (ACE2)), uma enzima que é encontrada na superfície de muitas células humanas. A ACE2 é especialmente abundante nos pulmões, sendo este o motivo pelo qual este é o órgão que mais sofre durante uma infecção de SARS-CoV.
Os principais gêneros dentro da família Coronaviridae são Alphacoronavirus, Betacoronavirus, Gammacoronavirus e Deltacoronavirus. A maioria das espécies conhecidas de Alpha– e i infectam morcegos, então é provável que o ancestral desses gêneros fosse originalmente um vírus específico de morcegos que mais tarde mutou e adquiriu a habilidade de infectar outras espécies. Todos os coronavírus que infectam humanos pertencem a estes dois gêneros e incluem, além do SARS-CoV, também o MERS-CoV (que causa a Síndrome Respiratória do Oriente Médio) e vários vírus que causam sintomas brandos semelhantes a um resfriado, como HCoV-HKU1, HCoV-NL63 e HCoV-229E. Espécies de Gammacoronavirus infectam principalmente aves, mas ao menos uma espécie, Coronavirus HKU15, causa diarreia em porcos. O gênero Deltacoronavirus inclui o coronavírus aviário (Avian coronavirus (IBV)), que causa bronquite infecciosa em aves, e o coronavírus da beluga SW1 (Beluga whale coronavirus SW1), o único coronavírus conhecido que infecta um mamífero marinho.
Avian coronavirus. As glicoproteínas em forma de bastão são claramente visíveis no envelope.
O genoma dos coronavírus possui cerca de 30 mil nucleotídeos, sendo um dos maiores genomas entre vírus de RNA. O único vírus de RNA conhecido que possui um genoma maior, com cerca de 41 mil nucleotídeos, foi descoberto em 2018 e infecta… adivinha… planárias! Chamado de “nidovírus das células secretoras de planária” (Planarian secretory cell nidovirus (PSCNV)), ele pertence à ordem Nidovirales, que inclui os coronavírus e muitos outros vírus de RNA, mas parece ter se divergido da maioria dos membros de Nidovirales muito, muito tempo atrás. Talvez eu fale mais desse vírus específico e as implicações de sua descoberta numa postagem futura.
Vamos concluir esta postagem com uma rápida revisão do que aprendemos sobre SARS-CoV-2, o “coronavírus”:
Ele é um vírus de RNA, ou seja, possui grande potencial de mutação e é capaz de criar cópias de si mesmo no citoplasma do hospedeiro, sendo quase um vírus autossuficiente;
Ele possui um capsídeo helicoidal em torno do RNA;
Ele possui um envelope derivado da membrana do retículo endoplasmático do hospedeiro, que é a razão pela qual pode ser morto tão facilmente com água e sabão;
Este envelope inclui grandes glicoproteínas em forma de bastão que o fazem parecer uma coroa solar em micrografias eletrônicas, de onde o nome coronavirus;
Ele é um membro do gênero Betacoronavirus, que inclui várias espécies conhecidas por infectarem morcegos, e esta é a razão pela qual sua origem numa sopa de morcego chinesa é muito provável.
Espero que esta postagem tenha ajudado você a ver mais desse vírus do que apenas sua habilidade de colapsar sociedades humanas.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências e leitura adicional:
Saberi A, Gulyaeva AA, Brubacher JL, Newmark PA, Gorbalenya AE (2018) A planarian nidovirus expands the limits of RNA genome size. PLoS Pathogens 14(11):e1007314. doi: 10.1371/journal.ppat.1007314
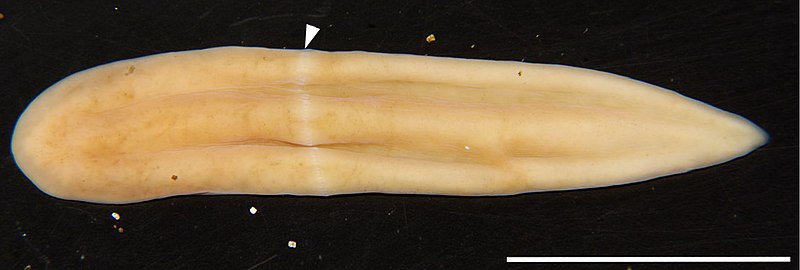
É hora de voltar aos fascinantes platelmintos e hoje decidi falar sobre uma das espécies mais bem estudadas neste grupo mesmo tendo sido apenas descrita formalmente 15 anos atrás. Seu nome é Macrostomum lignano, ou o macróstomo-de-Lignano.
Medindo 1 a 2 mm de comprimento, o macróstomo-de-Lignano pertence à ordem Macrostomida, um dos grupos mais basais de platelmintos. Seu corpo é alongado e transparente e há dois olhos pequenos perto da extremidade anterior, a qual possui um pequeno rostro (“focinho”). A boca é um pouco para trás do rostro. A extremidade posterior é larga, formando uma placa caudal com muitos órgãos adesivos arranjados em forma de U.
Morfologia básica do macróstomo-de-lignano. Extraído e traduzido de Lengerer et al. (2014).*
O macróstomo-de-Lignano foi coletado pela primeira vez em amostras marinhas na cidade de Lignano na costa do Mar Adriático no norte da Itália em 1995 e logo revelou ser bem adequado para culturas em laboratório. O ambiente natural desta espécie inclui a areia e outros sedimentos perto da costa. Ele evita a luz e, quando em repouso, permanece preso ao substrato pela placa caudal. Sua dieta inclui pequenos organismos, especialmente diatomáceas, as quais ingere usando sua faringe cilíndrica, similarmente a como a maioria dos platelmintos come.
Também como a maioria dos platelmintos, o macróstomo-de-Lignano e outros macrostômidos possuem células-tronco especiais chamadas neoblastos que preenchem seu corpo. Todas as células diferenciadas no corpo vêm de neoblastos e são continuamente substituídas por eles, visto que células diferenciadas não conseguem continuar reproduzindo. Os neoblastos também dão ao macróstomo-de-Lignano sua impressionante habilidade regenerativa como a de muitos outros platelmintos, como as planárias.
Mesmo antes de sua descrição formal em 2005, o macróstomo-de-Lignano já havia sido identificado como um potencialmente novo organismo modelo. Ele pode ser facilmente criado em laboratório em placas de Petri e alimentado com diatomáceas. Seu corpo possui cerca de 25000 células, o que é um número pequeno o bastante para facilitar estudos de desenvolvimento, regeneração, envelhecimento e expressão gênica e isso é exatamente o que vem sendo feito nas últimas décadas.
O macróstomo-de-Lignano é hermafrodita. Seu corpo contém dois testículos e dois ovários, e o aparelho copulador masculino contém um estilete, um órgão copulador enrijecido semelhante a um pênis. Quando dois macróstomos acasalam, eles tocam suas superfícies ventrais num estilo yin yang (assim como os carinhas da semana passada) e trocam esperma. Este comportamento é facilmente observado em laboratório e levou o macróstomo-de-Lignano a se tornar um organismo modelo para o estudo de seleção sexual também. Mas peraí! Seleção sexual num organismo hermafrodita? Sim! Eu discuti este tópico algum tempo atrás aqui.
Dois macróstomos-de-Lignano acasalando na posição yin yang. Foto de Lukas Schärer.***
Às vezes, quando dois macróstomos se encontram, eles não acham seu parceiro tão atraente, então ter seus ovos fertilizados por aquele cara não é de interesse de seu lado feminino. Contudo o lado masculino ainda é tão macho como qualquer outro e quer fertilizar tantos ovos quanto possível. Como resultado, se o parceiro não é bom o bastante, eles ainda o querem como fêmea mas não como macho. Mas o outro cara não está interessado em ser apenas fêmea, assim a cópula só ocorre se os dois parceiros aceitarem receber o esperma um do outro. “Eu deixo você fertilizar meus ovos se você me deixar fertilizar os seus.” Então é isso que eles fazem.
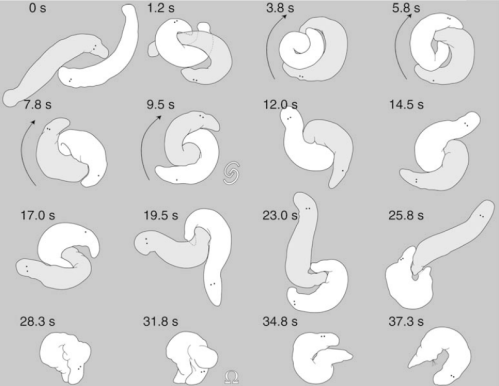
Um par de Macrostomum lignano acasalando. Veja como o branco, no final, se inclina sobre si mesmo e chupa o esperma do outroo cara para fora do poro feminino para se livrar dele. Note, no entanto, no último desenho, que os espermatozoides ainda estão presos ao poro feminino Não funcionou. Imagem extraída de Schärer et al. (2004).
Contudo depois de entregarem o esperma no corpo um do outro, eles se separam e podem nunca mais se ver de novo. Assim o lado feminino evoluiu uma estratégia para selecionar melhor o esperma. Quando o “parceiro ruim” se afasta, um macróstomo que recebeu esperma de baixa qualidade se curva sobre si mesmo, conecta a faringe ao poro genital feminino e chupa o esperma do outro cara para fora antes que ele tenha chance de fertilizar os ovos. Uma estratégia esperta, certo? Mas lembre-se: assim como este cara está tentando se livrar do esperma do outro, o outro pode estar fazendo o mesmo com o esperma deste. Assim uma estratégia precisa evoluir para prevenir a personalidade feminina de descartar o esperma. E isso é exatamente o que aconteceu! Os espermatozoides do macróstomo-de-Lignano possuem cerdas rígidas apontando para trás que, quando a célula é puxada para trás, entram nos tecidos do aparelho copulador feminino e ficam presos. Tentar puxar o esperma para fora é como tentar puxar espinhos de um porco-espinho para fora da pele.
Veja o comportamento em vídeo.
Agora o lado masculino recuperou a vantagem que o lado feminino teria se as cerdas não estivessem ali. Mas isso é a evolução, e seus efeitos em hermafroditas é como ter duas personalidades diferentes lutando uma contra a outra no mesmo corpo.
Precisa de ajuda para preparar seu artigo científico? Fale com a A1 Assessoria em Produção Acadêmica. Oferecemos serviços de tradução, revisão, formatação e preparação de figuras a preços acessíveis! Nosso e-mail: a1academica@gmail.com Se comentar que ouviu falar da A1 pelo Blog Natureza Terráquea, você ganha 10% de desconto na sua primeira tradução conosco!
– – –
Referências e leitura adicional:
Egger B, Ladurner P, Nimeth K, Gschwentner R, Rieger R (2006) The regeneration capacity of the flatworm Macrostomum lignano—on repeated regeneration, rejuvenation, and the minimal size needed for regeneration. Development Genes and Evolution 216:565–577. doi: 10.1007/s00427-006-0069-4
Ladurner P, Schärer L, Salvenmoser W, Rieger RM (2005) A new model organism among the lower Bilateria and the use of digital microscopy in taxonomy of meiobenthic Platyhelminthes: Macrostomum lignano, n. sp. (Rhabditophora, Macrostomorpha). Journal of Zoological Systematics and Evolutionary Research 43(2):114–126. doi: 10.1111/j.1439-0469.2005.00299.x
Lengerer B, Pjeta R, Wunderer J et al. (2014) Biological adhesion of the flatworm Macrostomum lignano relies on a duo-gland system and is mediated by a cell type-specific intermediate filament protein. Frontiers in Zoology 11:12. doi: 10.1186/1742-9994-11-12
Mouton S, Willems M, Braeckman BP, Egger B, Ladurner P, Schärer L, Borgonie G (2009) The free-living flatworm Macrostomum lignano: A new model organism for ageing research. Experimental Gerontology 44(4):243–249. doi: 10.1016/j.exger.2008.11.007
Pfister D, De Mulder K, Hartenstein V et al. (2008) Flatworm stem cells and the germ line: Developmental and evolutionary implications of macvasa expression in Macrostomum lignano. Developmental Biology 319(1):146–159. doi: 10.1016/j.ydbio.2008.02.045
Pfister D, De Mulder K, Philipp I et al. (2007) The exceptional stem cell system of Macrostomum lignano: Screening for gene expression and studying cell proliferation by hydroxyurea treatment and irradiation. Frontiers in Zoology 4:9. doi: 10.1186/1742-9994-4-9
Schärer L, Joss G, Sandner P (2004). Mating behaviour of the marine turbellarian Macrostomum sp.: these worms suck, Marine Biology145 (2) doi: 10.1007/s00227-004-1314-x
Wasik K, Gurtowski J, Zhou X et al. (2015) Genome and transcriptome of the regeneration-competent flatworm, Macrostomum lignano. PNAS 112(40):12462–12467. doi: 10.1073/pnas.1516718112